


Proteolytic and Biofilm-forming Capacity of Bacteria Isolated from a UHT Milk Processing Plant

Erika Molina1, Romina Camacho 1, Daniela Mejía 1, Martín Marcial-Coba 1*
Pontifical Catholic University of Ecuador, Quito 170143, Ecuador.
emolina1509@gmail.com, puyolromy197@gmail.com, Danylu.n.2000@gmail.com,
* Correspondence: msmarcial@puce.edu.ec; Tel.: (+593 989492886)
Pontifical Catholic University of Ecuador, Quito 170143, Ecuador.
emolina1509@gmail.com, puyolromy197@gmail.com, Danylu.n.2000@gmail.com,
* Correspondence: msmarcial@puce.edu.ec; Tel.: (+593 989492886)
ABSTRACT
The destabilization of milk during processing, caused by bacteria, represents a significant challenge for the dairy industry. This study aimed to determine the spoilage potential of 31 psychrotrophic, mesophilic, or thermophilic strains isolated from the UHT-milk production chain. This encompassed the biofilm-forming ability on polystyrene and stainless-steel surfaces, along with the strains' caseinolytic activity and heat resistance. The effect of 5 mM MgCl2 on the biofilm-forming capacity was also determined. The results indicated that the spoilage potential is highly strain-dependent. On polystyrene, Pseudomonas brenneri C10.3.4, Staphylococcus pasteuri RE1.9, and B. licheniformis SDP.15 were the strongest biofilm-formers of each group, whereas on stainless-steel, Pseudomonas lactis D1.11, Lacticaseibacillus paracasei P2.20 and Bacillus sonorensis C4.13 exhibited the highest capacity. Staphylococcus pasteuri RE1.9 and Staphylococcus epidermidis E3.6 biofilm cells showed the highest heat resistance.
Moreover, biofilm-born cells of the spore-forming B. licheniformis SDP.15 and B. sonorensis C4.13 were significantly more heat-resistant than their free-cell equivalents. Furthermore, 22 of the assessed strains showed proteolytic activity, although only the proteases produced by P. lactis E6.10 maintained their enzymatic activity after being treated at 92°C. These findings may contribute to developing strategies to enhance the dairy industry's long-term economic sustainability.
Keywords: UHT milk; spoilage potential; caseinolytic activity; biofilm-forming ability; thermal resistance
INTRODUCTION
Bovine milk is one of the most consumed foods worldwide due to its high nutritional content and energy contribution 1 . Due to these properties, milk constitutes a proper medium for the proliferation of multiple microorganisms. Therefore, microbial contamination throughout milk's industrial processing might lead to the proliferation of potential pathogenic or spoilage microorganisms in the commercial product 2 . The organisms found in raw dairy originate from different sources associated with cattle feed, cowshed environment, microbial contamination on the cow's udder's surface, and inadequate milking equipment cleaning3 . In addition, the sanitization of containers and the storage of physical-chemical conditions can also affect the microbial composition of raw milk 4 . Two main strategies have been widely applied in industrial milk processing to control or eliminate its microbial population 5 . In the first instance, raw milk is usually stored at refrigeration temperatures (4 - 7 °C) for a maximum of 48 hours to limit the growth of mesophilic and thermophilic bacteria 6 . Although this practice contributes to the temporary preservation of milk, it may also stimulate the growth of psychrotrophic microorganisms 7 . These are microorganisms adapted to grow at low temperatures (0 – 7 °C), even though their optimal and maximal growth temperatures are higher than 15 and 20 °C, respectively 8 .
Subsequently, as a second strategy, raw bovine milk undergoes industrial thermal processes to reduce or eliminate microbial load and to obtain a safe commercial product for human consumption 9 . Pasteurization involves milk exposure to a temperature of 72 °C for 15 s 10 . This aims to reduce the number of pathogenic bacterial cells to levels that do not represent a significant health hazard 11 . Likewise, in the ultra-high temperature (UHT) treatment, the heating of milk, between 135 to 150 °C for 1 to 4s, pursues a 9-log inactivation of its initial microbiota 12 . Combined with aseptic packaging, this process leads to the obtention of a commercially sterile product 13 .
Despite the efficiency of the UHT process, several studies have reported the isolation of multiple bacterial species in UHT milk samples, suggesting the occurrence of post-heat treatment contamination (e.g., container filling) or bacterial survival during thermal processing 14,15 . In this regard, several mechanisms can be associated with microbial resistance to heating steps during milk processing. For instance, bacterial endospores are dormant forms of vegetative cells resistant to adverse environmental conditions like starvation, bactericidal chemicals, and heat, among others 16 . Therefore, an elevated concentration of spore-forming bacteria in raw milk, particularly of highly heat-resistant strains, may increase these microbes' survival17 . Furthermore, most bacteria can form biofilms by adhering to multiple surfaces, such as stainless steel (SS), commonly used in the food industry 18 . A biofilm is a biologically active microbial community formed by cells of one or more species immersed in a matrix of exopolysaccharides 19 . The structure of this community creates an appropriate environment for microbial cells and is a self-protective mechanism against harsh conditions, including UV radiation, microbicidal compounds, and extreme pressure, pH, and temperature 20 . Therefore, this form of life enhances the survival of bacteria during heat treatments of milk, and its occurrence could constitute a source of cross-contamination in milk processing plants 21 . Moreover, the microorganisms in a biofilm could disperse in the food matrix immediately after contacting the contaminated surface 22 . This may compromise the quality and shelf-life of the product due to uncontrolled coagulation of the milk and the proliferation of bacteria resistant to thermal and chemical treatments 23 . Likewise, biofilms can generate hygiene inconveniences and financial impacts due to microorganisms catalyzing chemical and biological reactions that cause fouling in storage tanks and SS pipelines 24 .
In addition, certain extracellular enzymes produced by these bacteria can maintain their catalytic activity despite exposure to high temperatures 25 . Among them, lipases are lipolytic enzymes that break down lipids, including triglycerides in milk fat, into free fatty acids 26 . Proteases, on the other hand, degrade the major proteins present in milk, especially casein 27 . The destabilization of this protein by the action of proteolytic enzymes leads to changes in the organoleptic characteristics of the finished product, such as coagulation, gelation, and the formation of bitter flavors 28,29 .
Some conventional cleaning and disinfection processes are ineffective for the control of biofilm formation in dairy production and processing facilities. For this reason, various studies have focused on the suitability of potential biofilm inhibitory compounds. Recently, it has been reported that magnesium ions (Mg2+) affect the biofilm formation of Bacillus spp. through the downregulation of genes involved in the production of exopolysaccharides 30 . Therefore, utilizing Mg2+ ions may constitute an alternative strategy to improve the efficiency of sanitary practices in milk processing. Based on the background above, this research aimed to determine the spoilage potential of psychrotrophic, mesophilic, and thermophilic bacteria isolated from raw, pasteurized, and ultra-high temperature (UHT) processed milk by in vitro evaluation of their biofilm formation capacity, along with their response to 5mM magnesium chloride as an inhibitory agent. In addition, the proteolytic activity of each bacterial isolate was evaluated. The results obtained in this research could contribute to developing strategies focused on controlling bacterial contamination during milk processing and preventing commercial product spoilage.
MATERIAL AND METHODS
Bacterial strains
For this study, 14 psychrotrophic, 13 mesophilic, and 4 thermophilic strains were provided by the Food Microbiology Laboratory of the Microbiology career at PUCE. These strains were previously isolated from a UHT milk processing plant 31. Accession numbers of the 16S rDNA sequences of the evaluated strains are stated in the supplementary material.
The selected strains were reactivated on nutrient agar under the optimal temperature and growth time conditions for each strain to establish the collection of bacterial cultures used in the subsequent experiments. This was achieved using cultures previously preserved in BHI broth with 20% (v/v) glycerol.
Then, each bacterial isolate was propagated in 5 mL of sterile BHI broth under the same environmental conditions. Upon completion of incubation, an equal volume of glycerol, with a final concentration of 20%, was added, and the resulting suspension was divided into three aliquots of 1 mL, which were cryopreserved at -20 °C.
Screening of the biofilm-forming capacity in polystyrene microplates
The biofilm-forming capacity of the assessed strains was determined using 96-well flat-bottomed translucent polystyrene plates. Each well was loaded with 200 µL of sterile brain heart infusion (BHI) broth plus 20 µL of 24-hour liquid culture of each bacterial isolate. This suspension was incubated for 24 hours at 35 °C or 55 °C for mesophilic or thermophilic bacteria. Meanwhile, psychrotrophic strains were incubated at 7 °C for 10 days. The negative control consisted of individual aliquots of 220 µL sterile BHI broth dispensed in three independent wells.
After incubation, the content of the wells was discarded, and the plates were rinsed three times with Milli-Q water. Subsequently, the plates were allowed to dry at room temperature, and the biomass, adhered on the polystyrene surface, was fixed for 10 minutes by adding 220 µL of pure methanol into each well. After this period, methanol was poured off, and after a drying step at room temperature, a 220 µL volume of 0.05% crystal violet solution was added to each well. This step was followed by a 10-minute incubation period at room temperature. Right after this lapse, the dye was discarded, and the wells were rinsed with Milli-Q water. After drying, a 220 µL volume of an alcohol-acetone solution (50%-50% v/v) was added to each well, and the plates were incubated at room temperature for 15 minutes. Finally, this solution was transferred to a new polystyrene plate, and the optical density of each well was spectrophotometrically measured at a wavelength of 492 nm (OD492nm). The cut-off (CP) value was calculated through the sum of the mean OD492nm of three replicates of the negative control plus three standard deviations 18,32 .
Assessment of the biofilm-forming ability in the presence of 5 mM magnesium chloride as an inhibitory agent
Those strains that previously showed the ability to form biofilms (OD492nm > CP) were selected to evaluate their response to magnesium chloride as a biofilm inhibitory agent. For this purpose, the above-detailed methodology was followed, but a single variation involving the addition of magnesium chloride into the culture medium (final concentration = 5 mM) was incorporated into the experimental protocol 30 .
Biofilm-forming capacity on SS plates submerged in milk
This assay was performed according to the method described by Sadiq et al. 18 . Briefly, a 24-hour culture of biofilm-forming bacterial strains was individually inoculated into sterile glass flasks containing 100 mL of sterile skimmed milk. These suspensions were incubated for 24 hours at 30 °C for psychrotrophic strains, while mesophilic and thermophilic strains were incubated at 35 °C and 55 °C, respectively. Afterward, three sterile SS coupons (grade 316; 1 cm2; 2B finish) were immersed into the content of each of the flasks. In the case of mesophilic and thermophilic isolates, the cultures were further incubated for 24 hours, while psychrotrophic bacteria were incubated at 7 °C for 10 days. After incubation, the SS plates were aseptically removed and washed with sterile Milli-Q water. Subsequently, the SS coupons were placed in 50 ml conical tubes containing 10 ml of phosphate buffer solution (PBS) and 5 g of sterile glass beads to disperse the cells adhered on their surface. This preparation was stirred at 3000 rpm for 2 minutes, and subsequently, the contents of the conical tubes were diluted adequately in PBS. Thereafter, the dilutions were plated in triplicate on the surface of Tryptone Soy Agar (TSA) and incubated according to the optimal growth temperature of each bacterial group for 24 hours. At the end of this period, the formed colonies were quantified to determine the number of log UFC/mL and log UFC/cm2 developed in skimmed milk and on the surface of the SS plates, respectively.
Heat resistance of planktonic or biofilm cultures
Planktonic cultures in milk and biofilm cell suspensions of each bacterial isolate, obtained in the previous assay, were aliquoted in 1 mL fractions. These aliquots were thermally treated in a thermal block for 10 minutes at 80 °C. Non- and thermally treated suspensions were serially diluted in PBS. Subsequently, each dilution was inoculated in triplicate on the surface of TSA plates and incubated at the optimal growth temperature of each bacterium for 24 hours. Finally, bacterial colonies were counted to determine the number of viable log UFC/ml or log UFC/cm2 before or after the heat treatment 33 .
Screening of the proteolytic capacity of bacterial strains
The initially selected bacterial strains were individually, propagated in 5 ml of sterile BHI broth and incubated at their optimal growth temperature for 24 hours. After that, the cells were sedimented by centrifugation at 4000 rpm for 10 minutes 34 . The resulting supernatant was discarded, and under the same centrifugation parameters, the cells were washed with 2.5 ml of PBS. Subsequently, the sediment was suspended in the same volume of PBS, and its optical density, spectrophotometrically determined at a wavelength of 600 nm (OD600nm), was adjusted in the range of 0.5 to 0.8 by diluting each suspension in the required amount of PBS. Then, 25 µl of each bacterial suspension were inoculated in triplicate in perforated wells on a Plate count medium supplemented with skim milk 10%. The psychrotrophic strains were incubated at 7 °C for 10 days, while the mesophilic and thermophilic isolates were incubated for 48 h at 35 and 55 °C, respectively. After incubation, the formation of clarification halos around the inoculated wells was observed as a result of the proteolytic capacity of the bacterial isolates, and their diameters were measured.
Heat resistance of microbial extracellular proteases
The strains that showed proteolytic capacity in the previous assay were inoculated into conical tubes containing 10 ml of sterile BHI broth with skimmed milk 10% and incubated for 72 hours under the abovementioned environmental conditions. Thereafter, the cells were centrifuged at 6500 rpm for 30 minutes, and the supernatant was filtered and sterilized using a filter with a pore size of 0.22 µm. 35. Each cell-free supernatant (CFS) was evaluated at three pH levels. For this purpose, 200 µl aliquots of each CFS were individually mixed in a 1:1 ratio with 0.1 M sodium acetate buffer (pH 5.0), 0.1 M sodium phosphate buffer (pH 6.5), and 0.1 M Tris-HCl buffer (pH 8.0) 36 . In addition, the production of proteases was also evaluated without mixing the CFS with any buffer. Subsequently, to verify the presence of thermostable proteolytic enzymes, the supernatants were heat treated at 92 °C for 8 minutes and inoculated into perforated wells of milk agar. The untreated CFS was used as a positive control. After 24 hours of incubation, the formation of casein hydrolysis halos surrounding the inoculated well was observed, and their diameters were measured 37.
Inoculation effect of protease-producing bacteria in milk before sterilization
The bacterial strains that formerly showed proteolytic capacity were inoculated into 5 ml of sterile BHI broth and incubated for 24 hours at the optimal growth temperature for each strain until reaching an OD600nm corresponding to 0.5 - 0.8. Subsequently, 1 mL of a 1:100 dilution of each culture in PBS was inoculated into 5 ml of sterile skim milk. Afterwards, all bacterial suspensions were incubated at 4 °C for 48 hours. Thereafter, 1 ml of each homogenized sample was dispensed into sterile conical tubes, which were heat-treated at 130 °C for 30 seconds and immediately cooled down on ice. A sterility test was then conducted by seed inoculating 100 µl of each sample onto BHI agar. The tubes were incubated at 45 °C for 45 days. During this time, a daily visual inspection of each sample was performed to identify any effects on the milk, such as casein coagulation 34.
Statistical analysis
All experiments in this research were carried out in triplicate. The differences between the evaluated strains, corresponding to the mean of the OD492nm of biofilms developed on polystyrene, the concentration of planktonic or sessile cells in milk or the surface of SS, respectively, and the diameters of the casein digestion halos were determined through One-Way ANOVA. The post hoc Tukey multiple comparison test was used to analyze all possible strain pair combinations and ascertain whether the observed differences were statistically significant (P < 0.05). On the other hand, the Student's t-test was applied to compare the biofilm-forming capacity of each strain without MgCl2 against the OD492nm value obtained when cells of the same isolate were exposed to 5 mM MgCl2. Similarly, the same statistical analysis was used to compare the viability of planktonic or biofilm cells before and after thermal treatment. Significance was set at 0.05 for both statistical analyses.
RESULTS
Screening of the biofilm-forming capacity in polystyrene microplates
To evaluate the ability of biofilm formation, a cut-off point (CP) of 0.173 was established for the three analyzed bacterial groups. In the case of the psychrotrophic strains, it was observed that the OD492nm of 11 strains exceeded the CP value and, therefore, were considered biofilm-forming isolates. Pseudomonas brenneri C10.34 (OD492nm 0.818 ± 0.01) exhibited the highest biofilm-forming capacity. This was significantly higher (P < 0.05) than the OD492nm of all psychrotrophic strains.
For mesophilic bacteria, 12 out of 13 strains showed an OD492nm higher than that threshold. Staphylococcus pasteuri RE1.9 (OD492nm 0.337 ± 0.02) was the strain that demonstrated the highest biofilm-forming capacity (P < 0.05) compared to the absorbance of all strains. In the case of the thermophilic group, it was observed that three out of four isolates were capable of biofilm formation. This ability was significantly higher (P < 0.05) for Bacillus licheniformis SDP.15 (OD492nm 0.483 ± 0.05) when compared to the other two biofilm-forming strains.
Biofilm-forming ability in the presence of 5 mM magnesium chloride as an inhibitory agent
To assess the biofilm-forming ability of the isolates in the presence of 5 mM MgCl2, a unique CP was defined at 0.162. Of the 11 biofilm-forming psychrotrophic isolates, only Pseudomonas brenneri C10.34 and Pseudomonas fragi group E4.7 were inhibited since their OD492nm values were lower than the established CP. Additionally, it is essential to highlight that the absorbance, related to biofilm biomass, of every strain exposed to 5 mM MgCl2 was significantly lower than that developed in the absence of this compound, except for Pseudomonas fragi group C6.28, Pseudomonas fragi group E2.5, and Pseudomonas lactis D1.11 (Figure 1).

Figure. 1 OD492nm corresponding to the biofilm formation of psychrotrophic strains in the absence and presence of 5mM MgCl2
(*) Statistically significant reduction after treatment with 5mM MgCl2. A Strain showing a significantly higher biofilm-forming capacity in the absence of 5mM MgCl2 compared to all strains. B Strain showing a significantly higher biofilm-forming capacity in the presence of 5mM MgCl2 compared to all strains except C6.28, E11.31, and D1.11.
On the other hand, the biofilm-forming capacity of none of the mesophilic strains was inhibited by the presence of 5mM MgCl2, since their OD492nm was higher than the CP value. Although the absorbance of all the mesophilic strains, exposed to 5mM MgCl2, was lower than the value determined in the absence of this compound, a statistically significant reduction was only observed for Pantoea ananatis P.40, Pantoea conspicua D.23, Staphylococcus pasteuri RE1.9, Enterobacter hormaechei E8.16, Raoultella terrigena E7.12, and Lactococcus lactis subsp. lactis D.24 (Figure 2)

Figure. 2 OD492nm corresponding to the biofilm formation of mesophilic strains in the absence and presence of 5mM MgCl2
(*) Statistically significant difference after treatment with 5mM MgCl2. A Strain showing a significantly higher biofilm-forming capacity in the absence of 5mM MgCl2 compared to all strains. B Strain showing a significantly higher biofilm-forming capacity in the absence of 5mM MgCl2 compared to all strains.
(*) Statistically significant difference after treatment with 5mM MgCl2. A Strain showing a significantly higher biofilm-forming capacity in the absence of 5mM MgCl2 compared to all strains. B Strain showing a significantly higher biofilm-forming capacity in the absence of 5mM MgCl2 compared to all strains.
Finally, only the biofilm formation capacity of Enterococcus durans E1.4 was significantly reduced and inhibited by 5mM MgCl2 since its OD492nm value was lower than the established CP. Likewise, this capacity, compared to the previous treatment, was significantly reduced (P < 0.05) for Bacillus licheniformis SDP.15 when exposed to this compound (Figure 3).
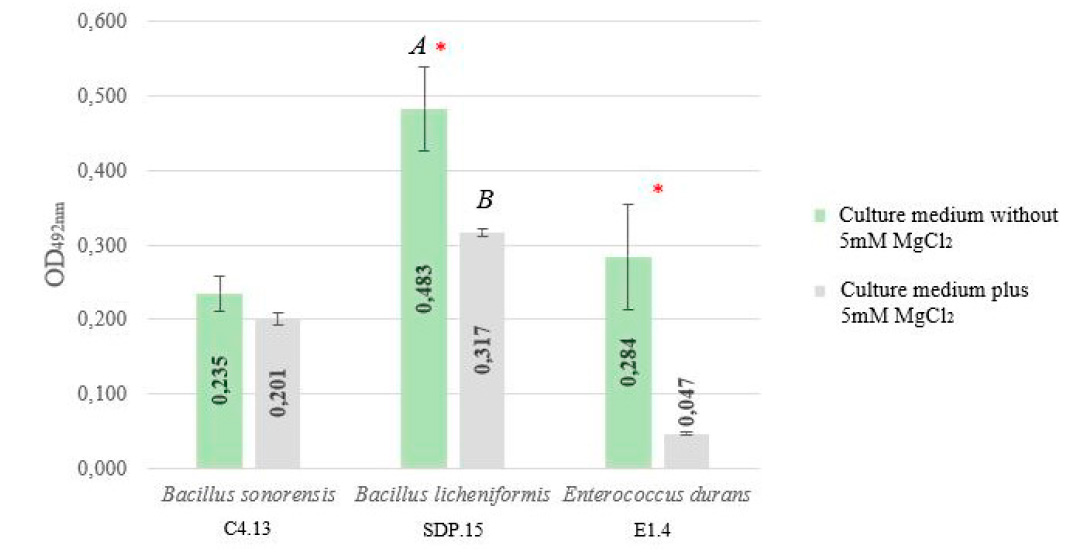
Figure. 3 OD492nm corresponding to the biofilm formation of thermophilic strains in the absence and presence of 5mM MgCl2
(*) Statistically significant difference after treatment with 5mM MgCl2. A Strain showing a significantly higher biofilm-forming capacity in the absence of 5mM MgCl2 compared to all strains. B Strain showing a significantly higher biofilm-forming capacity in the absence of 5 mM MgCl2 than all strains.
Biofilm-forming capacity on SS plates submerged in milk
In this study, the cell counts of biofilms recovered from SS plates were compared to the number of planktonic cells for each strain. Among the psychrotrophic bacteria group, it was observed that the highest biofilm cell concentration, equivalent to 8.56 ± 0.49 log CFU/cm2, corresponded to Pseudomonas lactis D1.11. In comparison to the rest of the psychrotrophic strains, this was statistically higher than the sessile cell counts of Pseudomonas fragi group E11.31, Pseudomonas lactis E6.10, Pseudomonas fragi group C6.28, Pseudomonas brenneri C10.34, Pseudomonas brenneri C7.29, Pseudomonas fragi group E2.5, Pseudomonas lurida C4.18, and Pseudomonas fragi group E4.7 (Table 1). In contrast, this latter strain exhibited the lowest biofilm-forming capacity on SS (6.86 ± 0.07 log CFU/cm2).
On the other hand, the planktonic cell concentration of all the psychrotrophic strains propagated in milk, was higher than 7 log CFU/mL. In this regard, it was observed that Pseudomonas fragi group C6.28 achieved the highest count (9.59 ± 0.03 log CFU/mL), which was significantly higher than all strains, except for Pseudomonas fragi group E11.31, Pseudomonas lactis E6.10, and Pseudomonas fragi group E4.7. However, the cells of Pseudomonas fragi group C6.28, recovered from the SS plates, exhibited the second lowest concentration (7.14 ± 0.01 log CFU/cm2) in this group. Additionally, Pseudomonas fragi group E2.5 was the strain that showed the lowest counts of planktonic cells (7.11
Likewise, Lacticaseibacillus paracasei subsp. tolerans P2.20 was the mesophilic isolate with the highest concentration (8.98 ± 0.01 log CFU/cm2) of biofilm cells attached to SS. This result was statistically higher than the counts obtained for all strains, except for Enterococcus durans E12.33 and Enterobacter hormaechei E8.16 (Table 1). Interestingly, L. paracasei subsp. tolerans P2.20 also showed the highest count of planktonic cells (8.96 ± 0.02 log CFU/mL) within this group. On the contrary, the cells of Dermacoccus nishinomiyaensis C1.1, adhered to SS plates, reached a concentration of 7.38 ± 0.01 log CFU/cm2. This count was significantly lower (P < 0.05) than all the analyzed strains.
Regarding the thermophilic strains, it was observed that Bacillus sonorensis C4.13 exhibited significantly higher counts of both sessile cells (8.23 ± 0.01 log CFU/cm2) and planktonic cells (8.83 ± 0.02 log CFU/mL) compared to all the other isolates. On the other hand, Bacillus licheniformis SDP.15 was the thermophilic strain that reflected the less efficient biofilm-forming capacity on SS, reaching a concentration equal to 5.25 ± 0.08 log CFU/cm2. For free cells, the lowest value was 8.03 ± 0.10 log CFU/mL for Enterococcus durans E1.4 (Table 1).
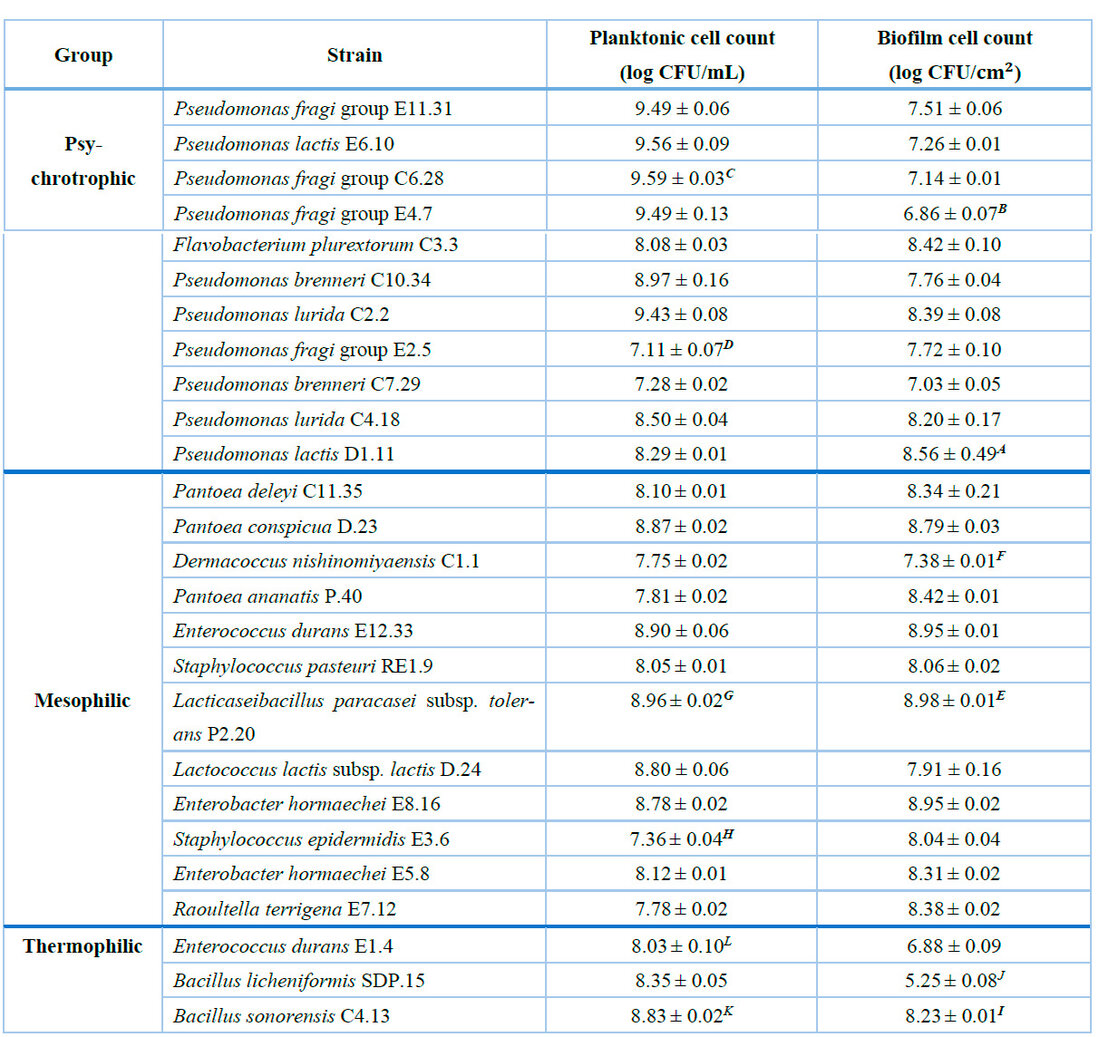
Planktonic cells were grown in sterile skimmed milk 10%, and biofilms were developed on the surface of stainless-steel plates submerged in the same matrix. The numerical values correspond to the mean count of three replicates ± the standard deviation of planktonic or biofilm cell concentration. A Biofilm cell count significantly higher (P < 0.05) than the concentration of all the psychrotrophic evaluated strains, except C3.3 and C2.2. B Biofilm cell count significantly lower compared to the count of all psychrotrophic isolates, except C6.28 and C7.29. C Planktonic cell count significantly higher than the concentration of C3.3, C10.34, C2.2, E2.5, C7.29, C4.18, and D1.11. D Planktonic cell count significantly lower than the concentration of all psychrotrophic strains. E Biofilm cell count significantly higher than the mean of all mesophilic isolates, except E12.33 and E8.16. F Biofilm cell count significantly lower than the concentration of all mesophilic strains. G Planktonic cell count significantly higher than the count of all mesophilic strains. H Planktonic cell count significantly lower than all concentrations of mesophilic isolates. I Biofilm cell count significantly higher compared to the count of all thermophilic strains. J Biofilm cell count significantly lower than the concentration of all thermophilic isolates. K Planktonic cell count significantly superior to the count of all thermophilic strains. L Planktonic cell count significantly lower than all thermophilic strains' mean.
Table 1. Concentration of planktonic cells grown in milk and biofilm bacteria developed on SS
Heat resistance of planktonic or biofilm bacterial cells
After the exposure of the psychrotrophic biofilm cells to thermal treatment, only Flavobacterium plurextorum C3.3, Pseudomonas lurida C2.2, and Pseudomonas lurida C4.18 showed detectable survival (> 3 log CFU/mL). The viability of these three strains suffered a reduction ranging from 3.57 ± 0.23 log CFU/mL for Flavobacterium plurextorum C3.3 to 4.10 ± 0.35 log CFU/mL for Pseudomonas lurida C4.18. These values were statistically lower (P < 0.05) compared to the reduction observed for the planktonic cells of each strain (Table 2). Therefore, the sessile cells of these strains might exhibit more resistance to high temperatures than their free-cell counterparts. Conversely, it was observed that the planktonic cells of Pseudomonas lactis D1.11, Pseudomonas fragi group C6.28, Pseudomonas fragi group E11.31, and Pseudomonas lactis E6.10 suffered a reduction between 3.19 ± 0.17 log CFU/mL to 5.32 ± 0.18 log CFU/mL. This result suggests that the free cells of these four strains exhibit more excellent resistance at that temperature (Table 2).
On the other hand, after the heat treatment, the biofilm cell viability, of all the evaluated mesophilic isolates, remained above 4.32 log CFU/mL (Table 2). During this assay, the viability loss oscillated between 0.03 ± 0.02 for Staphylococcus pasteuri RE1.9 and 3.59 ± 0.43 for Lactococcus lactis subsp. lactis D.24. The log reduction exhibited by sessile cells of Staphylococcus pasteuri RE1.9 was statistically lower than the viability loss of Pantoea conspicua D.23, Enterococcus durans E12.33, Lacticaseibacillus paracasei subsp. tolerans P2.20, Lactococcus lactis subsp. lactis D.24, and Enterobacter hormaechei E8.16. Similarly, planktonic cell survival of all the mesophilic strains, except for Pantoea deleyi C11.35, was evidenced throughout the heat treatment. However, the viability loss of every heat-resistant strain was significantly lower for cells in a sessile state than those in planktonic form. This finding suggests that biofilm cells of this bacterial group exhibit more excellent resistance compared to their planktonic counterparts (Table 2).
Moreover, the three thermophilic evaluated strains could survive the thermal treatment, either in a biofilm or planktonic state. Nevertheless, a biofilm cell reduction of 0.04 ± 0.01 and 1.76 ± 0.12 log CFU/mL for Bacillus sonorensis C4.13 and Bacillus licheniformis SDP.15, respectively, were significantly lower than the viability decreases experienced by planktonic cells of these strains (Table 2)
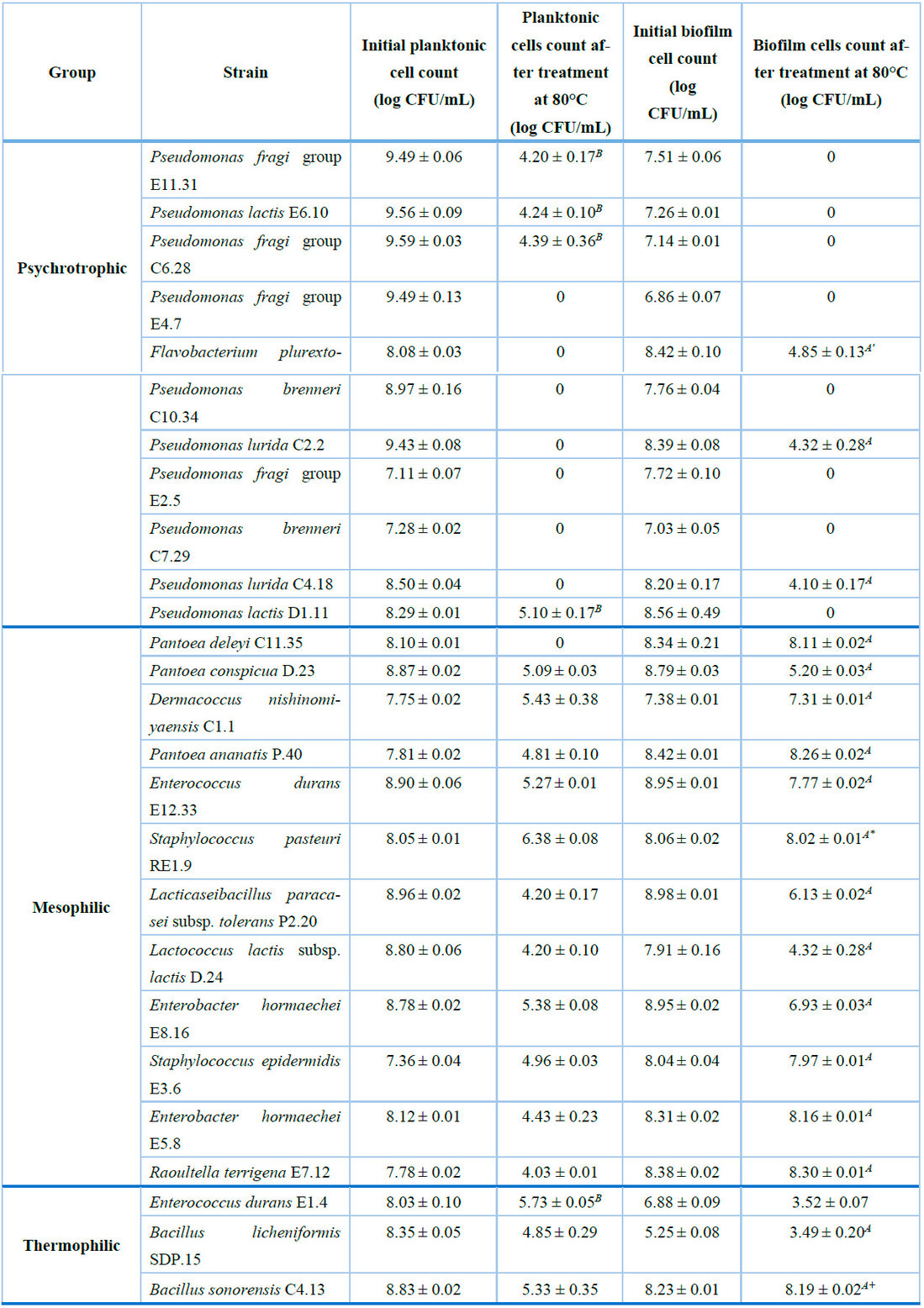
Each value corresponds to the mean count ± standard deviation of three replicates of planktonic or biofilm cells before and after the thermal treatment at 80°C. A Reduction of biofilm cells significantly lower (P < 0.05) than the reduction of planktonic cells after the thermal treatment. B Reduction of planktonic cells significantly lower than the reduction of biofilm cells after the thermal treatment. (') Strain that showed the lowest reduction of biofilm cells compared to all psychrotrophic isolates. (*) Mesophilic strain that showed the lowest reduction of sessile cells compared to D.23, E12.33, P2.20, D.24, and E8.16. (+) Strain that showed the lowest reduction of sessile cells compared to all thermophilic isolates.
Table 2. Heat resistance of planktonic or biofilm cells of psychrotrophic, mesophilic, or thermophilic isolates
Screening of protease-producing bacterial strains
As a result of their proteolytic activity, the growth of 22 out of 31 evaluated strains led to the formation of casein degradation halos around each inoculum (Table 3). Among psychrotrophic bacteria, eleven isolates exhibited halos whose diameters ranged from 1.1 ± 0.1 for Pseudomonas brenneri C10.34 to 3.2 ± 0.2 cm for Pseudomonas lurida C2.2. The latter strain showed a significantly higher (P < 0.05) caseinolytic capacity in comparison to the rest of the psychrotrophic strains. Furthermore, eight mesophilic strains showed halos whose diameters oscillated from 1.1 ± 0.1 to 2.9 ± 0.1 cm for Lacticaseibacillus paracasei subsp. tolerans P2.20 and Staphylococcus epidermidis E3.6, respectively. This latest isolate exhibited the highest proteolytic activity compared to the other strains within this group (Table 3).
On the other hand, three thermophilic strains, out of four, exhibited protease-producing capacity. Their digestion halos showed diameters ranging between 2.8 ± 0.4 cm for Bacillus sonorensis C4.13 and 3.3 ± 0.6 cm for Bacillus licheniformis SDP.15. On average, these three strains exhibited higher proteolytic activity compared to those belonging to the other two bacterial groups (Table 3).
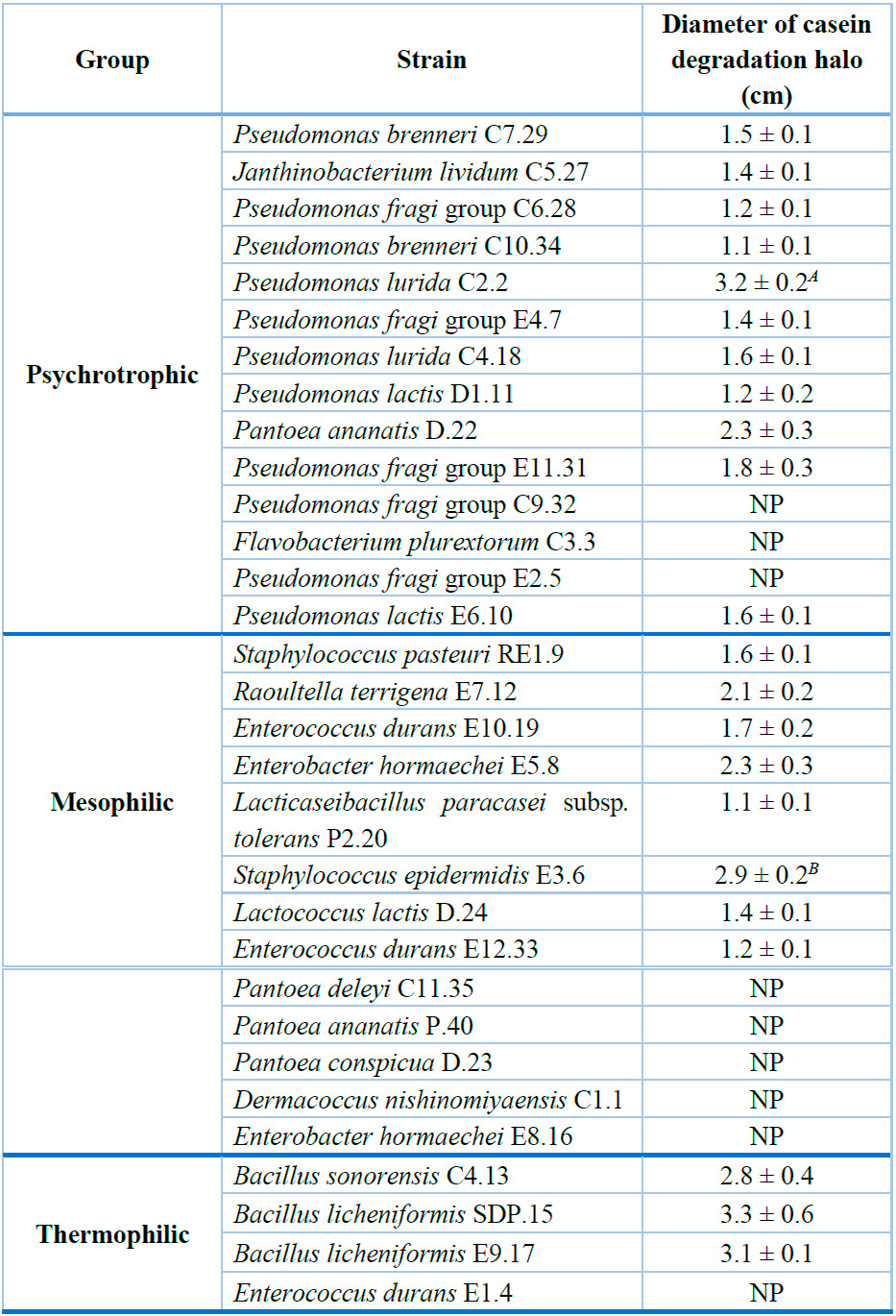
The numerical values correspond to the average of three replicates ± standard deviation of the diameter of casein digestion halos. A Strain that exhibited the highest proteolytic capacity compared to all psychrotrophic strains (P < 0.05). B Strain that showed the highest caseinolytic activity compared to all mesophilic strains. "NP" Indicates that it did not exhibit proteolytic capacity.
Table 3. Screening of the proteolytic capacity of psychrotrophic, mesophilic, and thermophilic bacterial isolates
Heat resistance of bacterial extracellular proteases
After thermally treating the CFS, individually obtained from the culture of protease-producing bacterial strains, it was evidenced that, uniquely, the inoculation of the supernatant extracted from the cultivation of Pseudomonas lactis E6.10, led to the development of casein degradation halos. Like the control without heat treatment, the diameter of clearing zones, around the inoculated wells, was 1.6 ± 0.1 cm. This was evident in the case of unbuffered CFS and when pH was adjusted to 5, 6.5, and 8.
Inoculation effect of protease-producing bacteria in milk before sterilization
Except for milk inoculated with Pseudomonas brenneri C7.29, Pseudomonas fragi group C6.28, Pseudomonas brenneri C10.34, Pseudomonas lactis D1.11, or Pseudomonas fragi group E11.31, most of the samples, previously contaminated with individual protease-producing strains and subsequently sterilized, suffered syneresis during storage at 45 °C. This phenomenon was evidenced by the casein coagulation and its subsequent separation from the whey. This effect was noticeable during the initial seven days for samples containing remaining metabolites of psychrotrophic and thermophilic bacteria. On the other hand, changes were observed in milk containing metabolic products of mesophilic bacteria only after 30 days of incubation (Table 4)
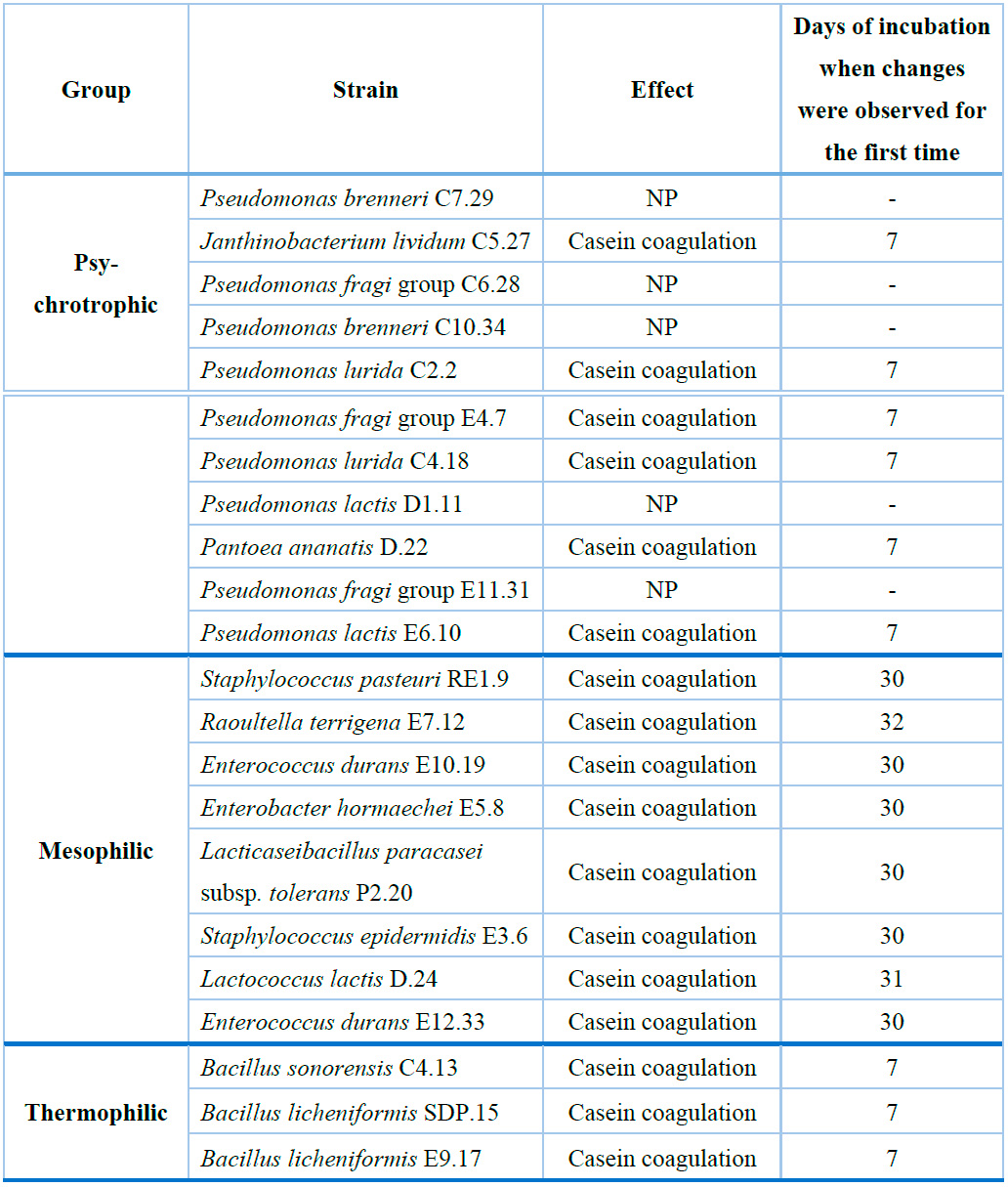
NP: No effect on sterile milk after 45 days of incubation at 45°C. (-) No changes were observed on any day.
Table 4. Effect of inoculation of psychrotrophic, mesophilic, and thermophilic protease-producing bacteria in milk before sterilization
DISCUSSION
This study revealed that Gram-negative psychrotrophic Pseudomonas brenneri C10.34 exhibited the highest biofilm-forming capacity on polystyrene wells. In this connection, it has previously been described that multiple milk-originated strains belonging to Pseudomonas spp., including P. fluorescens and P. brenneri, displayed an efficient biofilm-forming capacity, on polystyrene plates 38–40 . Additionally, consistent with our outcomes, Rossi et al. 39 have documented that an extended incubation time (≥ 48h) at a low temperature (10 °C), stimulates biofilms development of P. fluorescens and P. brenneri strains. This finding also suggests that this temperature enhances motility and, consequently, the initial adhesion of bacteria cells to the substrate and the occurrence of homogeneous properties of their exopolysaccharides. Similarly, Gram-positive thermophilic Bacillus licheniformis SDP15 and mesophilic Staphylococcus pasteuri RE1.9, along with Gram-negative mesophilic Raoultella terrigena E7.12 showed a relatively high biofilm-forming ability on the same substratum. This capacity of the mentioned milk-contaminant bacteria has been well described in other investigations. For example, several strains of B. licheniformis have been reported as robust biofilm-formers, particularly when isolated from dairy production environments 41 . In the same context, milk isolates of coagulase-negative staphylococci, which include S. pasteuri, possess key factors, mainly polysaccharide intercellular adhesin (poly-N-acetylglucosamine), that allow this genus to form robust biofilms on multiple surfaces 42-44 . Moreover, in the present investigation, it was observed that biofilm-forming ability was highly variant, even among strains of the same species. For instance, the OD492nm of P. brenneri C10.34 was approx. 4 times higher than that of P. brenneri C7.29. In like manner, Pseudomonas lurida C2.2 displayed a more robust biofilm-forming capacity (OD492nm 0.513 ± 0.1) than that of Pseudomonas lurida C4.18 (OD492nm 0.198 ± 0.005). This phenomenon has also been observed in different studies, suggesting that the ability to form biofilms is strain-dependent and closely tied to the adaptive potential to new environmental conditions 39,45 . This intra-species variability relies on multiple aspects like the adhesion ability, related to cell surface hydrophobicity and the development of appendages (pili, curli, fimbriae, flagella) 46 . Factors determining biofilm maturation (e.g., type and concentration of produced EPS plus transport mechanisms through the cell membrane) are also associated with strain-dependent biofilm-forming capacity 47 .
On the other hand, various strategies have been explored in the food industry to control biofilm formation, encompassing different bacterial species or strains. Among these strategies, using MgCl2 as an inhibiting agent stands out as a potential alternative. Despite the crucial role those divalent cations like Mg2+ and Ca2+ play in the physiology of bacterial cells, an excessive amount is detrimental 48 . It has been demonstrated that these ions can influence the electrostatic interactions and surface adhesion processes, dependent on bacterial physiology, by acting as cellular cations and enzymatic cofactors 49 . Furthermore, it has also been demonstrated that Mg2+ ions affect biofilm formation by negatively regulating the expression of extracellular matrix genes 50 . Thus, in the present study, we assessed the effect of 5 mM MgCl2 on the bacterial isolate's biofilm-forming capacity on 96-well polystyrene plates. This treatment repressed the biofilm formation (OD492nm < CP) of Pseudomonas brenneri C10.34, Pseudomonas fragi group E4.7, and Enterococcus durans E1.4. Furthermore, in response to 5mM MgCl2 exposure, the OD492nm associated with the adhered biomass of 13 out of 23 remaining strains was significantly reduced (P < 0.05), although their biofilm-forming ability was not inhibited. Between the abovementioned isolates, the OD492nm of Bacillus licheniformis SDP.15, Pantoea ananatis P.40, Staphylococcus pasteuri RE1.9, Pseudomonas lurida C2.2, Pseudomonas brenneri C10.34, and Enterococcus durans E1.4 (ascendingly ordered), was reduced by approx.1.5 to 6 times when exposed to the inhibiting compound. Consistent with our outcomes, Mulcahy and Lewenza 51 have reported that Mg2+ limitation triggers the biofilm formation of Pseudomonas aeruginosa strains, which is closely related to the Pseudomonas species included in our investigation. Similarly, it has been reported that the biofilm biomass of Enterobacter cloacae BF-17 was significantly reduced as a response to 0.5 – 256 mM MgCl2.6H2O exposure 52 . Moreover, concerning the effect of this compound on Bacillus spp. biofilm formation, variable results have been described. For instance, Ben-Ishay et. al. 30 observed that MgCl2, in concentrations equal to or higher than 5 mM, is detrimental to biofilm formation of Bacillus spp. strains, while Oknin, Steinberg and Shemesh 50 suggest that this effect only occurs on Bacillus subtilis when exposed to at least 25 mM MgCl2. This may also explain that in contrast to Bacillus licheniformis SDP.15, biofilms of Bacillus sonorensis C4.13 were not significantly altered during this treatment. Along with the latter, in this study biofilm formation ability was not changed for the other nine strains, including Pseudomonas fragi group C6.28 and E2.5, Pseudomonas lactis D1.11, Enterobacter hormaechei E5.8 and C1.1, Enterococcus durans E12.33, and Staphylococcus epidermidis E3.6. This suggests that 5 mM MgCl2 might not be effective regarding biofilm inhibition for the latter strains. In this connection, it has been proposed that 16 mM Mg2+ stimulates biofilm formation on plastic surfaces instead of inhibiting 53. In addition, based on the results as mentioned earlier, it was noted that the strains evaluated in the present investigation, even belonging to the same species, responded variably to 5 mM MgCl2 exposure. Consistent with this, other studies suggest that the biofilm-inhibiting effect of Mg2+ may also be a concentration, species, or strain-dependent property 54 . Based on this, in further studies, the response of our isolates to MgCl2 in a concentration range from 5 to 50 mM shall be evaluated to determine the most efficient value in terms of the biofilm inhibitory capacity of this compound.
Moreover, it was also observed that all bacterial isolates could grow in milk, reaching concentrations exceeding 7 log CFU/mL in a planktonic state. Nevertheless, robust growth in this matrix did not necessarily lead to a high concentration of cells adhered to SS plates. For example, Pseudomonas fragi group E4.7, Enterococcus durans E1.4, and Bacillus licheniformis SDP. 15 reached a concentration lower than 7 log CFU/cm2 adhered to SS, whereas Pseudomonas lactis D1.11, Lacticaseibacillus paracasei subsp. tolerans P2.20 and Bacillus sonorensis C4.13 were the strains that exhibited the highest concentrations (> 8 logs CFU/cm2) of attached cells on the same surface. In this connection, it has been settled that biofilm formation, in the presence of milk, results from the interaction of several abiotic and biotic factors. These involve the proportion of metal ions in surface composition (Ni, Fe, Mn, among others), its hydrophobicity, a solution pH close to the isoelectric point of bacteria, and electrostatic attraction between cells and the metal surfaces 55,56 . Additionally, raw milk proteins may reduce the bacterial ability to form biofilms on SS 57 , whereas the occurrence of milk surfactant compounds and heat-denatured protein deposits stimulates this capacity 59 . At the same time, bacterial adhesion on SS is also a strain-dependent property and compromises the formation of S-layer, EPS production, and the presence of filamentous appendages 56 . In addition, it has been outlined that butyric acid in milk is a critical factor that triggers B. licheniformis biofilm formation 58 and that SS provides better support than polystyrene for biofilm development of this species 18,60 . Nevertheless, the present research results do not directly support that claim since B. licheniformis SDP. 15 exhibited a high biofilm-forming ability on polystyrene but was the less efficient biofilm-former strain on SS in contact with milk. This finding aligns with the outcomes of Parkar et al. 61 who observed that the adhesion of both spores and vegetative cells of B. licheniformis was reduced when SS plates were immersed in 1% and 10% skimmed milk. Furthermore, it has been demonstrated that the nature of this effect appears to vary depending on the organism, substrate, and milk proteins 62
On the other hand, Lacticaseibacillus paracasei subsp. tolerans P2.20 was one of the weakest biofilm formers strains on polystyrene but the strongest on SS. Consistently, Dat et al. 63 have suggested that Lacticaseibacillus paracasei is a strong biofilm former on SS in the presence of milk. The precipitation of milk proteins might also stimulate this phenomenon due to bacterial fermentation and a pH decrease to 5.5 63 . In contrast, it has been demonstrated that UHT milk (whole, semi-skimmed, and skimmed) significantly alters the hydrophobicity of steel, regardless of contact time 64,65 . This alteration promotes the pre-adsorption of α, β, and κ-casein and results in a 20% reduction in Staphylococcus aureus adhesion compared to clean steel. It is presumed that this effect could be similar in other species of this genus 62 . Our results support those findings since S. pasteuri RE1.9 showed the fourth lowest count of biofilm cells on SS plates immersed in skimmed milk, while on polystyrene, it proved the most efficient.
As a proof-of-concept, biofilm-originated cells of all the mesophilic strains along with psychrotrophic Flavobacterium plurextorum C3.3, Pseudomonas lurida C2.2, and C4.18; when exposed to heat treatment (80 °C for 10 min), exhibited a significantly lower viability loss compared to their planktonic counterparts. As a function of several mechanisms, the heat-stress response of biofilms is generally more effective than that of free planktonic cells 66,67 . In the same context, the log reduction suffered by the sessile cells of the thermophilic isolates Bacillus licheniformis SDP.15 and Bacillus sonorensis C4.13, after heat treatment, was 1.98 and 87.5 times, respectively, lower than the viability loss of the corresponding bacteria in the planktonic state. Our results suggest that the heat resistance might be enhanced when bacteria of those strains are biofilm-originated, although a remarkable survival was expected due to their intrinsic capacity to produce heat-resistant spores. In this connection, Simmonds et al. 68 demonstrated that spores of B. licheniformis, B. cereus, and Geobacillus stearothermophilus adhered to SS were significantly more resistant (P < 0.05) to high-temperature exposure (90°C, 95°C, 100°C, and 110°C) than non-biofilm-originated spores. Similarly, Huang, Flint and Palmer 33 , have described that the D90°C values of biofilm spores are significantly extended by at least 6 minutes compared to the D90°C corresponding to free-floating spores. This might occur due to additional protection against adverse environments provided by the polymeric matrix surrounding the endospores embedded in a biofilm 41,69 .
Surprisingly, an insignificant viability loss of biofilm-originated cells of Staphylococcus pasteuri RE1.9 and S. epidermidis E3.6 resulted from exposure to high temperature. In line with this, a study conducted by Almatroudi et al. 70 reported that biofilm cells of S. aureus experienced a 2 log reduction when exposed to 121°C for 20 minutes, whereas planktonic cells suffered a viability loss higher than 8 log. This remarkable resistance might be attributed to the elevated proportion of anteiso-C19 fatty acids formerly described as the membrane of staphylococcal biofilm cells 71 . The high melting point of these molecules leads to a reduction in membrane fluidity and consequently enhances resistance to thermal stress 71 . In addition, according to a study conducted by Montanari et al. 72 , this great thermal resistance is present in other species of this genus, and thus, it may support our findings.
On the other hand, the concentration of thermally treated viable planktonic cells of Pseudomonas fragi group E11.31, C6.28, along with Pseudomonas lactis E6.10 and D1.11, remained close to 4 log CFU/mL. In contrast, biofilm cells suffered a loss of viability below the assay's detection limit after heat treatment. In this connection, most strains within this group exhibited an initial concentration of sessile cells close to 7 log CFU/mL, whereas planktonic cells exceeded 8 log CFU/mL. This observation suggests that the higher initial concentration of planktonic cells influenced the resulting survival above the assay's detection limit despite biofilm cells' commonly associated heat resistance. Moreover, it should be considered that biofilm cells of Pseudomonas lurida C4.18 and C2.2, closely related to the above strains, suffered an average cell death of approximately. 4 log CFU/mL from an initial concentration of 8.20 ± 0.17 log CFU/mL and 8.39 ± 0.08 log CFU/mL, respectively. In a hypothetical scenario in which sessile cells of Pseudomonas fragi group E11.31 and C6.28 plus Pseudomonas lactis E6.10, with initial concentrations close to 7 logs CFU/mL, suffered the same log reduction, a non-detectable survival might be observed as it resulted in the present study. This notable observation might be reinforced by the pattern observed for Pseudomonas fragi group E2.5 and Pseudomonas brenneri C7.29, whose initial populations of both cell types were approx. 7 log CFU/mL and, consistently, no survival was detected. In line with this, it has been observed that high population density might be associated with enhanced cell heat resistance, owing to the accumulation of protective compounds against thermal stress (e.g., trehalose and catalase), in addition to the synthesis of heat shock proteins 73,74 . Therefore, it is logical to expect that cells with lower concentrations would be more susceptible to adverse conditions, thus supporting the findings of this study.
On the other hand, Enterococcus durans E1.4 exhibited a significantly higher initial concentration of planktonic cells (8.03 ± 0.10 log CFU/mL) than the concentration of its counterparts recovered from biofilms (6.88 ± 0.09 log CFU/mL). It is noteworthy that, despite biofilm cells' relatively low initial cell density, a survival of at least 3 log CFU/mL was detected after thermal treatment. However, this value was lower when compared to that of free-living cells. Overall, it has been widely described that Enterococcus species possess high thermal resistance attributed to genetic adaptations that stabilize cellular structures or may even repair the damage caused by heat 75,76 . The characteristic peptidoglycan-rich cell wall of these Gram-positive bacteria provides greater rigidity and protection 77 . Therefore, it is well known that this type of bacteria may exhibit more excellent resistance compared to Gram-negative species. These advantages enable them to maintain cell viability throughout thermal treatments of up to 80°C, consistent with the outcomes observed in the present investigation.
In contrast, E. durans E12.33, classified in this study as a mesophilic growth strain, showed a remarkable heat resistance since they suffered a viability loss corresponding to 3.63 ± 0.07 and 1.18 ± 0.03 log CFU/mL for planktonic and sessile cells, respectively, after an initial concentration close to 9 log CFU/mL of both cell states, was thermally treated. In this connection, our findings align with those reported by McAuley et al. 76 , who previously highlighted the significant variability regarding heat resistance among enterococci strains. This variability could result from a complex interplay of factors, including regulating gene expression in the biosynthetic pathways of spermidine and putrescine 75 . These polyamines play a pivotal role in bacterial survival, as they possess the ability to stabilize DNA structure and uphold cellular membrane integrity under thermal stress conditions 78 .
In addition, 22 out of 31 analyzed strains exhibited proteolytic capacity, as evidenced by the formation of digestion halos around the inoculated wells in milk agar. Within this context, a notable variation in caseinolytic potential was observed among closely related strains. For example, two out of the five strains belonging to the fragi group of Pseudomonas were not capable of casein degradation. This discrepancy may be attributed to species diversity within this taxonomic group, which includes P. fragi, P. lundensis, P. taetrolens, and P. psychrophila 79,80 . Interestingly, this variability was also evident among strains of the same species, as demonstrated in the case of P. lurida C2.2, which exhibited significantly larger diameters of casein degradation halos than all other psychrotrophic isolates, including P. lurida C4.18. In line with this Caminero et al. 81 , have suggested that the type and proportion of proteases vary significantly among different taxonomic groups and strains. Consistently, Ribeiro Júnior et al. 82 reported the variability, in terms of proteolytic ability, of different Pseudomonas spp. isolates in bovine milk might be related to its capacity to produce the AprX metalloprotease, which catalyzes the hydrolysis of α, β, and κ-casein 83 . This enzyme is codified by the aprX gene, which, in the genome of Pseudomonas spp., is part of the aprX-lipA operon. In this connection, it has been reported that the operon structure is variable among species or strains of this genus 84,85 since this operon contains genes related to secretion mechanisms (aprD, aprE, aprF, prtAB), a protease inhibitor (aprI), and a lipase (lipA) 84 . Consequently, Maier et al. 84 demonstrated that the strain-specific proteolytic capacity relies on the genetic structure of the aprX-lipA operon. Besides these intrinsic traits, environmental parameters like temperature 83 , calcium, and iron content 85-87 influence the casein digestion ability 88 .
Furthermore, significant variability of the caseinolytic potential was observed among the Enterococcus durans strains evaluated in this study. At the end of the incubation period, only the two mesophilic growth isolates displayed hydrolysis zones on the culture medium, whereas the thermophilic strains exhibited no proteolytic ability. Within this context, it has been documented that the production of gelatinase (GelE), an extracellular zinc metalloprotease, and serine protease (SprE), which form part of the proteolytic system of enterococci strains isolated from dairy products, are not representative of all species of this genus 89-93 . The respective encoding genes, gelE, and sprE, are co-transcribed within the same operon 92,94 . Variability of the enzymatic activity may be attributed to the fact that these genes are only expressed when the enterococci population has reached high cell densities and are thus under the control of a regulatory fsr quorum sensing (QS) system consisting of three regulatory genes (fsrA, fsrB, and fsrC) 91,94,95 . In this regard, Pessione et al. 92 have suggested that modifying these regulatory genes might impair protease functionality. This could account for the absence of proteolytic activity.
In addition, it was evident that, on average, Bacillus spp. strains exhibited notably more giant degradation halos than those in the other two evaluated groups. Previously, it has been reported that the genus Bacillus can produce large amounts of enzymes such as alkaline serine proteases and zinc metalloproteases, which represent over 95% of their extracellular proteolytic activity 96,97 . Within this context, Mehta et al. 98 observed that the enzymes produced by this genus can cleave β and κ-casein. Likewise, previous studies have documented that the proteolytic potential of B. licheniformis might vary as a result of the incubation temperature 100 . In this connection, and consistent with our results, Sayem, Alam and Hoq 101 reported an optimal temperature range between 37°C and 50°C for the activity of B. licheniformis proteases, and consequently, extensive casein hydrolysis zones were observed when cultivated in skim milk agar medium (SMA).
Moreover, it has been outlined that most heat-resistant proteases related to UHT milk spoilage are represented by the AprX metalloprotease produced by species belonging to Pseudomonas spp. 25,102 . Nevertheless, in this research, only the CFS, heat-treated at 92°C, of Pseudomonas lactis E6.10 could form casein hydrolysis zones when inoculated into plate count agar supplemented with skim milk 10%. Although the aprX gene is widespread among Pseudomonas spp., its nucleotidic sequence shows variability within different strains of this genus 28 . This affects the flexibility of the protein's three-dimensional structure and, consequently, its ability to accurately refold after exposure to high temperatures 103 . This aspect and the aprX gene's expression level could elucidate why heat-resistant proteases were not observed for P. lactis D1.11 or the rest of the Pseudomonas spp. isolates evaluated in this study. As previously mentioned, this residual enzymatic activity is one of the primary factors contributing to the deterioration of UHT dairy products 104–106 . Therefore, in future research, it would be pertinent to extract and characterize this protease and its thermo-stability at 135°C to determine its potential impact during the storage of the final product, which shall be assessed.
Although the heat resistance of proteases was demonstrated only for those produced by Pseudomonas lactis E6.10, when bacterial strains were individually inoculated into milk 48 h before its sterilization, casein coagulation, associated with bacterial metabolism, was observed in 17 out of 22 sterile milk samples throughout 45 days of storage. This outcome might be linked to a pH decrease in milk due to the production of bacterial metabolites, mainly organic acids 107 . Previous investigations have highlighted that casein reaches its isoelectric point at a pH of 4.6 leading to increased compaction of its micellar structure 108,109 . Therefore, our findings suggest that casein coagulation, observed in this assay, and synaeresis were likely caused by the bacterial metabolism before the sterilization process rather than protease activity.
Milk-spoilage-related microorganisms represent critical challenges for the food industry. In our study, a remarkable biofilm-forming capacity on both polystyrene and SS surfaces, as well as heat resistance of sessile cells and proteolytic ability, was exhibited by several strains. This includes isolates like S. pasteuri RE1.9, S. epidermidis E3.6, E. durans E12.33, Lacticaseibacillus paracasei subsp. tolerans P2.20, B. sonorensis C4.13, B. licheniformis SDP.15, P. lurida C2.2, and Raoultella terrigena E7.12. In this connection, the presence of bacteria, showing such spoilage potential, in a UHT milk production plant represents a considerable hazard that could affect the quality and safety of dairy products and the operational efficiency of related industries. Likewise, the control of thermostable protease production, such as those secreted by P. lactis E6.10, should be further studied since they also challenge the stability of the commercial product during its shelf life.
On the other hand, the properties exhibited by Lacticaseibacillus paracasei subsp. tolerans P2.20, Lactococcus lactis D.24, and Bacillus licheniformis SDP.15 shall be considered within the investigation of their putative biotechnological applications. This may involve their probiotic potential, the acidification rate of lactic acid bacteria strains during milk fermentation, and the production of industrial-relevant metabolites, among others.
This research provides a comprehensive insight into the variability of biofilm-forming capacity, caseinolytic activity, and heat resistance among bacterial strains isolated from the production chain of UHT milk. Thus, these findings may contribute to developing strategies to enhance the efficiency of processes like milk collection and transportation, thermal treatment, packaging, and industrial storage. We emphasize the need for pilot studies to confirm the applicability of our findings in practical settings. Additionally, it is essential to conduct further research to elucidate the underlying mechanisms of microbial spoilage. Finally, developing and assessing new control strategies for managing spoilage microorganisms is necessary. These integrated approaches are crucial for enhancing the dairy industry's safety, integrity, and long-term sustainability.
CONCLUSION
This research suggests that the milk spoilage potential of bacteria could be strain-dependent, showing significant variations even between different strains of the same species. Among the 31 evaluated isolates, 26 could form biofilms on polystyrene. In this connection, Pseudomonas brenneri C10.34 was the strongest psychrotrophic biofilm-former on polystyrene surfaces, whereas, on the same substratum, Staphylococcus pasteuri RE1.9 and Bacillus licheniformis SDP.15 were the most efficient biofilm-producers among mesophilic and thermophilic strains, respectively. Moreover, it was observed that a concentration of 5 mM MgCl2 significantly reduced this capacity for 16 strains, suggesting that the utilization of this compound, after further investigation of adequate concentrations, could be efficient in terms of biofilm control. However, the biofilm biomass of isolates like Lacticaseibacillus paracasei subsp. tolerans P2.20, among others, were not affected when exposed to 5 mM MgCl2, although it did not behave as a strong biofilm-former on polystyrene. The latter mesophilic bacterium, the psychrotrophic isolate P. lactis D1.11, and the thermophilic culture of B. sonorensis C4.13 exhibited the highest capacity of biofilm development on SS in each of the evaluated groups. On the other hand, when exposed to high temperatures, the sessile cells of 17 strains were substantially more resistant than their planktonic counterparts.
Interestingly, among the psychrotrophic isolates, only cells recovered from biofilms of Flavobacterium plurextorum C3.3, P. lurida C2.2, and P. lurida C4.18 maintained their viability after the heat treatment. Similarly, although the heat-resistance of bacterial endospores has been widely described, biofilm-born cells of the spore-forming strains B. licheniformis SDP.15 and B. sonorensis C4.13 were significantly more resistant to high temperatures than their free cells equivalents. However, the biofilm cells of S. pasteuri RE1.9 and S. epidermidis E3.6 proved to be the most heat-resistant of the entire experiment.
Moreover, although 22 of the assessed strains secreted extracellular proteases, only those produced by P. lactis E6.10 maintained their enzymatic activity after being treated at 92°C. This research highlights the critical need for optimized strategies involving the control of bacterial contamination during UHT milk production based on the determination of strain-dependent unique characteristics in terms of their spoilage potential. This approach would mitigate the risks associated with biofilm-forming, heat-resistant, and proteolytic bacteria and could ensure that processes in a dairy production plant remain robust and efficient. Moreover, the observed properties of some of the evaluated strains should be considered for further studies related to the potential biotechnological applications of these microorganisms or their metabolites.
Supplementary Material: Accession numbers of the 16S rDNA sequences of the evaluated strains are stated in the supplementary material.
Author Contributions: Erika Molina: Writing – original draft; Investigation; Writing – review & editing; Formal analysis; Data curation; Methodology. Romina Camacho: Methodology. Daniela Mejía: Methodology. Martín Marcial-Coba: Conceptualization; Funding acquisition; Writing – review & editing; Investigation; Supervision; Methodology; Formal analysis.
Funding: Pontifical Catholic University of Ecuador funded this research.
Conflicts of Interest: The authors declare no conflict of interest.
REFERENCES
1. Fuentes-Coto G, Ruiz-Romero RA, Sánchez-Gómez JI, Ávila-Ramírez DN, Escutia-Sánchez J. ANALYSIS OF ORGANIC MILK : DESIRABLE ATTRIBUTES FOR ITS TRANSFORMATION. 2021.
2. Lu M, Wang NS. Spoilage of Milk and Dairy Products. In: The Microbiological Quality of Food: Foodborne Spoilers. Elsevier Inc.; 2017. p. 151–78.
3. Deddefo A, Mamo G, Asfaw M, Amenu K. Factors affecting the microbiological quality and con-tamination of farm bulk milk by Staphylococcus aureus in dairy farms in Asella, Ethiopia. BMC Mi-crobiol. 2023 Dec 1;23(1).
4. Decimo M, Morandi S, Silvetti T, Brasca M. Characterization of Gram-negative psychrotrophic bac-teria isolated from Italian bulk tank milk. J Food Sci. 2014 Oct 1;79(10):M2081–90.
5. Machado SG, Baglinière F, Marchand S, Coillie E Van, Vanetti MCD, Block J De, et al. The biodi-versity of the microbiota producing heat-resistant enzymes responsible for spoilage in processed bovine milk and dairy products. Front Microbiol. 2017;8(MAR):1–22.
6. Yuan L, Sadiq FA, Burmølle M, Wang N, He G. Insights into psychrotrophic bacteria in raw milk: A review. J Food Prot. 2019 Jul 1;82(7):1148–59.
7. Decimo M, Morandi S, Silvetti T, Brasca M. Characterization of Gram-negative psychrotrophic bac-teria isolated from Italian bulk tank milk. J Food Sci. 2014 Oct 1;79(10):M2081–90.
8. de Oliveira GB, Favarin L, Luchese RH, McIntosh D. Psychrotrophic bacteria in milk: How much do we really know? Vol. 46, Brazilian Journal of Microbiology. Sociedade Brasileira de Microbiologia; 2015. p. 313–21.
9. Fatih M, Barnett MPG, Gillies NA, Milan AM. Heat Treatment of Milk: A Rapid Review of the Im-pacts on Postprandial Protein and Lipid Kinetics in Human Adults. Front Nutr. 2021 Apr 30;8.
10. Dash KK, Fayaz U, Dar AH, Shams R, Manzoor S, Sundarsingh A, et al. A comprehensive review on heat treatments and related impact on the quality and microbial safety of milk and milk-based products. Food Chemistry Advances. 2022 Oct;1:100041.
11. Macdonald LE, Brett J, Kelton D, Majowicz SE, Snedeker K, Sargeant JM. A systematic review and meta-analysis of the effects of pasteurization on milk vitamins, and evidence for raw milk consumption and other health-related outcomes. Vol. 74, Journal of Food Protection. 2011. p. 1814–32.
12. Melini F, Melini V, Luziatelli F, Ruzzi M. Raw and heat-treated milk: From public health risks to nutritional quality. Beverages. 2017;3(4):1–33.
13. Barbosa-Cánovas G V, Kelly AL, Larsen LB. Food Engineering Series Series Editor: Agents of Change Enzymes in Milk and Dairy Products. In: Food Engineering Series [Internet]. 2021. p. 315–50. Available from: http://www.springer.com/series/5996
14. Kmiha S, Aouadhi C, Klibi A, Jouini A, Béjaoui A, Mejri S, et al. Seasonal and regional occurrence of heat-resistant spore-forming bacteria in the course of ultra-high temperature milk production in Tunisia. J Dairy Sci. 2017 Aug 1;100(8):6090–9.
15. Scheldeman P, Herman L, Foster S, Heyndrickx M. Bacillus sporothermodurans and other highly heat-resistant spore formers in milk. In: Journal of Applied Microbiology. 2006. p. 542–55.
16. Stoeckel M, Lücking G, Ehling-Schulz M, Atamer Z, Hinrichs J. Bacterial spores isolated from in-gredients, intermediate and final products obtained from dairies: thermal resistance in milk. Dairy Sci Technol. 2016 Jul 1;96(4):569–77.
17. Ohkubo Y, Uchida K, Motoshima H, Katano N. Microbiological safety of UHT milk treated at 120 °C for 2 s, as estimated from the distribution of high-heat-resistant Bacillus cereus in dairy environments. Int Dairy J. 2019 Apr 1;91:36–40.
18. Sadiq FA, Flint S, Yuan L, Li Y, Liu TJ, He GQ. Propensity for biofilm formation by aerobic meso-philic and thermophilic spore forming bacteria isolated from Chinese milk powders. Int J Food Mi-crobiol. 2017 Dec 4;262:89–98.
19. Navia DP, Samuel Villada H, Mosquera SA. BIOFILMS IN THE FOOD INDUSTRY BIOFILMES NA INDÚSTRIA DE ALIMENTOS. 2010.
20. Yin W, Wang Y, Liu L, He J. Biofilms: The microbial "protective clothing" in extreme environments. Int J Mol Sci. 2019 Jul 2;20(14).
21. Huang Y, Flint SH, Palmer JS. The heat resistance of spores from biofilms of Bacillus cereus grown in tryptic soy broth and milk. Int Dairy J. 2021 Dec 1;123.
22. Marchand S, De Block J, De Jonghe V, Coorevits A, Heyndrickx M, Herman L. Biofilm Formation in Milk Production and Processing Environments; Influence on Milk Quality and Safety. Vol. 11, Comprehensive Reviews in Food Science and Food Safety. 2012. p. 133–47.
23. Teh KH, Flint S, Brooks J, Knight G. Biofilms in the Dairy Industry, First Edition. Edited. 2015.
24. González F, Fontecha P, Rodríguez J. Biofilms: Contaminacion Cruzada En Industria Alimentaria. 2015;
25. Stoeckel M, Lidolt M, Stressler T, Fischer L, Wenning M, Hinrichs J. Heat stability of indigenous milk plasmin and proteases from Pseudomonas: A challenge in the production of ultra-high temperature milk products. Vol. 61, International Dairy Journal. Elsevier Ltd; 2016. p. 250–61.
26. Márquez C, Piramanrique K, Carrascal AK, Clavijo B, Quevedo B. Determinación cuantitativa de proteasas de bacterias psicrotróficas aisladas de leche cruda.
27. Zhang C, Boeren S, Zhao L, Bijl E, Hettinga K. The Impact of Low-Temperature Inactivation of Protease AprX from Pseudomonas on Its Proteolytic Capacity and Specificity: A Peptidomic Study. Dairy. 2023 Mar 1;4(1):150–66.
28. Matéos A, Guyard-Nicodème M, Baglinière F, Jardin J, Gaucheron F, Dary A, et al. Proteolysis of milk proteins by AprX, an extracellular protease identified in Pseudomonas LBSA1 isolated from bulk raw milk, and implications for the stability of UHT milk. Int Dairy J. 2015 Oct 1;49:78–88.
29. Chavan RS, Chavan SR, Khedkar CD, Jana AH. UHT milk processing and effect of plasmin activity on shelf life: A review. Vol. 10, Comprehensive Reviews in Food Science and Food Safety. 2011. p. 251–68.
30. Ben-Ishay N, Oknin H, Steinberg D, Berkovich Z, Reifen R, Shemesh M. BRIEF COMMUNICA-TION Enrichment of milk with magnesium provides healthier and safer dairy products. NPJ Biofilms Microbiomes [Internet]. 2017;3:24. Available from: www.nature.com/npjbiofilms
31. Chávez N. Detección de bacterias formadoras de esporas en leche vacuna cruda, pasteurizada y pro-cesada por Ultra Alta Temperatura (UHT) [tesis de grado]. Quito: Pontificia Universidad Católica del Ecuador; 2023. Disponible en: https://repositorio.puce.edu.ec/items/c2dfe037-5f26-4bb3-a917-f0642d31e24d
32. Alonso VPP, de Oliveira Morais J, Kabuki DY. Incidence of Bacillus cereus, Bacillus sporother-modurans and Geobacillus stearothermophilus in ultra-high temperature milk and biofilm formation capacity of isolates. Int J Food Microbiol. 2021;354(February).
33. Huang Y, Flint SH, Palmer JS. The heat resistance of spores from biofilms of Bacillus cereus grown in tryptic soy broth and milk. Int Dairy J. 2021 Dec 1;123.
34. Morandi S, Pica V, Masotti F, Cattaneo S, Brasca M, De Noni I, et al. Proteolytic traits of psy-chrotrophic bacteria potentially causative of sterilized milk instability: Genotypic, phenotypic and peptidomic insight. Foods. 2021 May 1;10(5).
35. Ouertani A, Chaabouni I, Mosbah A, Long J, Barakat M, Mansuelle P, et al. Two new secreted pro-teases generate a casein-derived antimicrobial peptide in Bacillus cereus food born isolate leading to bacterial competition in milk. Front Microbiol. 2018 Jun 4;9(JUN).
36. Lim YH, Foo HL, Loh TC, Mohamad R, Abdullah N. Comparative studies of versatile extracellular proteolytic activities of lactic acid bacteria and their potential for extracellular amino acid productions as feed supplements. J Anim Sci Biotechnol. 2019 Mar 7;10(1).
37. Stuknyte M, Decimo M, Colzani M, Silvetti T, Brasca M, Cattaneo S, et al. Extracellular thermostable proteolytic activity of the milk spoilage bacterium Pseudomonas fluorescens PS19 on bovine caseins. J Dairy Sci. 2016 Jun 1;99(6):4188–95.
38. Lauer Cruz K, de Souza da Motta A. Characterization of biofilm production by Pseudomonas fluo-rescens isolated from refrigerated raw buffalo milk. J Food Sci Technol. 2019 Oct 1;56(10):4595–604.
39. Rossi C, Serio A, Chaves-López C, Anniballi F, Auricchio B, Goffredo E, et al. Biofilm formation, pigment production and motility in Pseudomonas spp. isolated from the dairy industry. Food Control. 2018 Apr 1;86:241–8.
40. Zarei M, Rahimi S, Saris PEJ, Yousefvand A. Pseudomonas fluorescens group bacterial strains interact differently with pathogens during dual-species biofilm formation on stainless steel surfaces in milk. Front Microbiol. 2022 Oct 26;13.
41. Ostrov I, Paz T, Shemesh M. Robust biofilm-forming bacillus isolates from the dairy environment demonstrate an enhanced resistance to cleaning-in-place procedures. Foods. 2019 Apr 1;8(4).
42. Otto M, Gov MN. Staphylococcal Biofilms. National Institutes of Health [Internet]. 2008;322:207–28. Available from: http://www.cdc.gov/ncidod/hip/NNIS/2004NNISreport.pdf
43. Tremblay YDN, Caron V, Blondeau A, Messier S, Jacques M. Biofilm formation by coagulase-negative staphylococci: Impact on the efficacy of antimicrobials and disinfectants commonly used on dairy farms. Vet Microbiol. 2014 Aug 27;172(3–4):511–8.
44. De Visscher A, Piepers S, Haesebrouck F, Supré K, De Vliegher S. Coagulase-negative Staphylococcus species in bulk milk: Prevalence, distribution, and associated subgroup- and species-specific risk factors. J Dairy Sci. 2017 Jan 1;100(1):629–42.
45. Rossi C, Chaves-López C, Serio A, Goffredo E, Goga BC, Paparella A. Influence of incubation con-ditions on biofilm formation by pseudomonas fluorescens isolated from dairy products and dairy manufacturing plants. Ital J Food Saf. 2016;5(3).
46. Lianou A, Nychas GJE, Koutsoumanis KP. Strain variability in biofilm formation: A food safety and quality perspective. Food Research International. 2020 Nov 1;137.
47. Ksontini H, Kachouri F, Hamdi M. Dairy Biofilm: Impact of Microbial Community on Raw Milk Quality. J Food Qual. 2013 Aug;36(4):282–90.
48. Oknin H, Steinberg D, Shemesh M. Magnesium ions mitigate biofilm formation of Bacillus species via downregulation of matrix genes expression. Front Microbiol. 2015;6(SEP).
49. Demishtein K, Reifen R, Shemesh M. Antimicrobial properties of magnesium open opportunities to develop healthier food. Nutrients. 2019 Oct 1;11(10).
50. Oknin H, Steinberg D, Shemesh M. Magnesium ions mitigate biofilm formation of Bacillus species via downregulation of matrix genes expression. Front Microbiol. 2015;6(SEP).
51. Mulcahy H, Lewenza S. Magnesium limitation is an environmental trigger of the pseudomonas aeru-ginosa biofilm lifestyle. PLoS One. 2011;6(8).
52. Zhou G, Li L jie, Shi Q shan, Ouyang Y sheng, Chen Y ben, Hu W feng. Efficacy of metal ions and isothiazolones in inhibiting enterobacter cloacae BF-17 biofilm formation. Can J Microbiol. 2014 Jan;60(1):5–14.
53. Dunne WM, Burd EM. The Effects of Magnesium, Calcium, EDTA, and pH on the In Vitro Adhesion of Staphylococcus epidermidis to Plastic. Microbiol Immunol. 1992;36(10):1019–27.
54. Wang T, Flint S, Palmer J. Magnesium and calcium ions: roles in bacterial cell attachment and biofilm structure maturation. Biofouling. 2019 Oct 21;35(9):959–74.
55. Jindal S, Anand S, Metzger L, Amamcharla J. Short communication: A comparison of biofilm de-velopment on stainless steel and modified-surface plate heat exchangers during a 17-h milk pasteuri-zation run. J Dairy Sci. 2018 Apr 1;101(4):2921–6.
56. Dula S, Ajayeoba TA, Ijabadeniyi OA. Bacterial biofilm formation on stainless steel in the food pro-cessing environment and its health implications. Vol. 66, Folia Microbiologica. Springer Science and Business Media BV; 2021. p. 293–302.
57. Al-Makhlafi H, Nasir A, McGuire J, Daeschel M. Adhesion of Listeria monocytogenes to silica surfaces after sequential and competitive adsorption of bovine serum albumin and beta-lactoglobulin. Appl Environ Microbiol. 1995 May;61(5):2013-5.
58. Pasvolsky R, Zakin V, Ostrova I, Shemesh M. Butyric acid released during milk lipolysis triggers biofilm formation of Bacillus species. Int J Food Microbiol. 2014 Jul 2;181:19–27.
59. Marchand S, De Block J, De Jonghe V, Coorevits A, Heyndrickx M, Herman L. Biofilm Formation in Milk Production and Processing Environments; Influence on Milk Quality and Safety. Vol. 11, Comprehensive Reviews in Food Science and Food Safety. 2012. p. 133–47.
60. Md Zain SN, Bennett R, Flint S. The Potential Source of B. licheniformis Contamination During Whey Protein Concentrate 80 Manufacture. J Food Sci. 2017 Mar 1;82(3):751–6.
61. Parkar S, Flint S, Palmer J, Brooks J. Factors influencing attachment of thermophilic bacilli to stainless steel. J Appl Microbiol. 2001;901–8.
62. Barnes LM, Lo MF, Adams MR, Chamberlain AHL. Effect of Milk Proteins on Adhesion of Bacteria to Stainless Steel Surfaces Downloaded from [Internet]. Vol. 65, APPLIED AND ENVIRONMENTAL MICROBIOLOGY. 1999. Available from: http://aem.asm.org/
63. Dat NM, Hamanaka D, Tanaka F, Uchino T. Control of milk pH reduces biofilm formation of Bacillus licheniformis and Lactobacillus paracasei on stainless steel. Food Control. 2012 Jan;23(1):215–20.
64. Hamadi F, Asserne F, Elabed S, Bensouda S, Mabrouki M, Latrache H. Adhesion of Staphylococcus aureus on stainless steel treated with three types of milk. Food Control. 2014 Apr;38(1):104–8.
65. Hamadi F, Latrache H, Asserne F, Elabed S, Zahir H, Saad IK, et al. Quantitative Adhesion of Staphylococcus aureus on Stainless Steel Coated with Milk. Food Nutr Sci. 2013;04(03):299–304.
66. Mikkelsen H, Duck Z, Lilley KS, Welch M. Interrelationships between colonies, biofilms, and planktonic cells of Pseudomonas aeruginosa. J Bacteriol. 2007 Mar;189(6):2411–6.
67. Pawluk AM, Kim D, Jin YH, Jeong KCC, Mah JH. Biofilm-associated heat resistance of Bacillus cereus spores in vitro and in a food model, Cheonggukjang jjigae. Int J Food Microbiol. 2022 Feb 16;363.
68. Simmonds P, Mossel BL, Intaraphan T, Deeth HC. Heat Resistance of Bacillus Spores When Adhered to Stainless Steel and Its Relationship to Spore Hydrophobicity. Vol. 66, Journal of Food Protection. 2003.
69. Postollec F, Mathot AG, Bernard M, Divanac'h ML, Pavan S, Sohier D. Tracking spore-forming bacteria in food: From natural biodiversity to selection by processes. Int J Food Microbiol. 2012 Aug 1;158(1):1–8.
70. Almatroudi A, Tahir S, Hu H, Chowdhury D, Gosbell IB, Jensen SO, et al. Staphylococcus aureus dry-surface biofilms are more resistant to heat treatment than traditional hydrated biofilms. Journal of Hospital Infection. 2018 Feb 1;98(2):161–7.
71. Abdallah M, Chataigne G, Ferreira-Theret P, Benoliel C, Drider D, Dhulster P, et al. Effect of growth temperature, surface type and incubation time on the resistance of Staphylococcus aureus biofilms to disinfectants. Appl Microbiol Biotechnol. 2014;98(6):2597–607.
72. Montanari C, Serrazanetti DI, Felis G, Torriani S, Tabanelli G, Lanciotti R, et al. New insights in thermal resistance of staphylococcal strains belonging to the species Staphylococcus epidermidis, Staphylococcus lugdunensis and Staphylococcus aureus. Food Control. 2015 Apr 1;50:605–12.
73. Liu X, Ng C, Ferenci T. Global Adaptations Resulting from High Population Densities in Escherichia coli Cultures [Internet]. Vol. 182, JOURNAL OF BACTERIOLOGY. 2000. Available from: https://journals.asm.org/journal/jb
74. Mah TFC, O'toole GA. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol [Internet]. 2001;9(1):34. Available from: http://tim.trends.com0966
75. Wang L, Qiao L, Li A, Chen L, He B, Liu G, et al. Integrative Multiomics Analysis of the Heat Stress Response of Enterococcus faecium. Biomolecules. 2023 Mar 1;13(3).
76. McAuley CM, Gobius KS, Britz ML, Craven HM. Heat resistance of thermoduric enterococci isolated from milk. Int J Food Microbiol. 2012 Mar 15;154(3):162–8.
77. Silhavy TJ, Kahne D, Walker S. The bacterial cell envelope. Vol. 2, Cold Spring Harbor perspectives in biology. 2010.
78. Hussain T, Tan B, Ren W, Rahu N, Dad R, Kalhoro DH, et al. Polyamines: therapeutic perspectives in oxidative stress and inflammatory diseases. Vol. 49, Amino Acids. Springer-Verlag Wien; 2017. p. 1457–68.
79. Yumoto I, Kusano T, Shingyo T, Nodasaka Y, Matsuyama H, Okuyama H. Assignment of Pseudo-monas sp. strain E-3 to Pseudomonas psychrophila sp. nov., a new facultatively psychrophilic bacterium. Extremophiles. 2001;5(5):343–9.
80. Garrido-Sanz D, Meier-Kolthoff JP, Göker M, Martín M, Rivilla R, Redondo-Nieto M. Genomic and genetic diversity within the Pseudomonas fluoresces complex. PLoS One. 2016 Feb 1;11(2).
81. Caminero A, Guzman M, Libertucci J, Lomax AE. The emerging roles of bacterial proteases in intes-tinal diseases. Vol. 15, Gut Microbes. Taylor and Francis Ltd.; 2023.
82. Ribeiro Júnior JC, de Oliveira AM, Silva F de G, Tamanini R, de Oliveira ALM, Beloti V. The main spoilage-related psychrotrophic bacteria in refrigerated raw milk. J Dairy Sci. 2018 Jan 1;101(1):75–83.
83. Zhang C, Bijl E, Hettinga K. Destabilization of UHT milk by protease AprX from Pseudomonas flu-orescens and plasmin. Food Chem. 2018 Oct 15;263:127–34.
84. Maier C, Hofmann K, Huptas C, Scherer S, Wenning M, Lücking G. Simultaneous quantification of the most common and proteolytic Pseudomonas species in raw milk by multiplex qPCR. Appl Microbiol Biotechnol [Internet]. 2020; Available from: https://doi.org/10.1007/s00253-021-11109-0
85. Woods RG, Burger M, Beven CA, Beacham IR. The aprX-lipA operon of Pseudomonas fluorescens B52 : a molecular analysis of metalloprotease and lipase production [Internet]. Vol. 147. 2015. Available from: www.microbiologyresearch.org
86. Maunsell B, Adams C, O'Gara F. Complex regulation of AprA metalloprotease in Pseudomonas flu-orescens M114: Evidence for the involvement of iron, the ECF sigma factor, PbrA and pseudobactin M114 siderophore. Microbiology (N Y). 2006 Jan;152(1):29–42.
87. Martins ML, Pinto UM, Riedel K, Vanetti MCD. Milk-deteriorating exoenzymes from Pseudomonas fluorescens 041 isolated from refrigerated raw milk. Brazilian Journal of Microbiology. 2015;46(1):207–17.
88. Aguilera-Toro M, Kragh ML, Thomasen AV, Piccini V, Rauh V, Xiao Y, et al. Proteolytic activity and heat resistance of the protease AprX from Pseudomonas in relation to genotypic characteristics. Int J Food Microbiol. 2023 Apr 16;391–393.
89. Anderson AC, Jonas D, Huber I, Karygianni L, Wölber J, Hellwig E, et al. Enterococcus faecalis from food, clinical specimens, and oral sites: Prevalence of virulence factors in association with biofilm formation. Front Microbiol. 2016 Jan 11;6(JAN).
90. Medeiros AW, Pereira RI, Oliveira D V, Martins PD, D’azevedo PA, Van Der Sand S, et al. Molecular detection of virulence factors among food and clinical Enterococcus faecalis strains in South Brazil [Internet]. 2014. Available from: www.sbmicrobiologia.org.br
91. Qin X, Singh K V, Weinstock GM, Murray BE. Effects of Enterococcus faecalis fsr Genes on Production of Gelatinase and a Serine Protease and Virulence Downloaded from [Internet]. Vol. 68, INFECTION AND IMMUNITY. 2000. Available from: http://iai.asm.org/
92. Pessione A, Lamberti C, Cocolin L, Campolongo S, Grunau A, Giubergia S, et al. Different protein expression profiles in cheese and clinical isolates of Enterococcus faecalis revealed by proteomic analysis. Proteomics. 2012 Feb;12(3):431–47.
93. Terzić-Vidojević A, Veljović K, Popović N, Tolinački M, Golić N. Enterococci from raw-milk cheeses: Current knowledge on safety, technological, and probiotic concerns. Vol. 10, Foods. MDPI; 2021.
94. Sifri CD, Mylonakis E, Singh K V., Qin X, Garsin DA, Murray BE, et al. Virulence effect of Enter-ococcus faecalis protease genes and the quorum-sensing locus fsr in Caenorhabditis elegans and mice. Infect Immun. 2002 Oct;70(10):5647–50.
95. Pillai SK, Sakoulas G, Gold HS, Wennersten C, Eliopoulos GM, Moellering RC, et al. Prevalence of the fsr locus in Enterococcus faecalis infections. J Clin Microbiol. 2002;40(7):2651–2.
96. Connelly MB, Young GM, Sloma A. Extracellular proteolytic activity plays a central role in swarming motility in Bacillus subtilis. J Bacteriol. 2004 Jul;186(13):4159–67.
97. Schallmey M, Singh A, Ward OP. Developments in the use of Bacillus species for industrial production. Vol. 50, Canadian Journal of Microbiology. 2004. p. 1–17.
98. Mehta DS, Metzger LE, Hassan AN, Nelson BK, Patel HA. The ability of spore formers to degrade milk proteins, fat, phospholipids, common stabilizers, and exopolysaccharides. J Dairy Sci. 2019 Dec 1;102(12):10799–813.
99. Sethi R. Optimization studies of Protease enzyme in in-vitro conditions from Bacillus licheniformis [Internet]. Vol. 3, International Research Journal of Biological Sciences. 2014. Available from: www.isca.me
100. Ribeiro Júnior JC, Tamanini R, de Oliveira ALM, Alfieri AA, Beloti V. Genetic diversity of ther-moduric spoilage microorganisms of milk from Brazilian dairy farms. J Dairy Sci. 2018 Aug 1;101(8):6927–36.
101. Sayem SMA, Alam J, Hoq MM. Effect Of Temperature, pH and Metal Ions On The Activity and Stability Of Alkaline Protease From Novel Bacillus Licheniformis MZK03 [Internet]. 2006. Available from: https://www.researchgate.net/publication/317031037
102. Baur C, Krewinkel M, Kutzli I, Kranz B, von Neubeck M, Huptas C, et al. Isolation and characteri-sation of a heat-resistant peptidase from Pseudomonas panacis withstanding general UHT processes. Int Dairy J. 2015 Oct 1;49:46–55.
103. Zhang C, Bijl E, Svensson B, Hettinga K. The Extracellular Protease AprX from Pseudomonas and its Spoilage Potential for UHT Milk: A Review. Compr Rev Food Sci Food Saf. 2019 Jul 1;18(4):834–52.
104. Chavan RS, Chavan SR, Khedkar CD, Jana AH. UHT milk processing and effect of plasmin activity on shelf life: A review. Vol. 10, Comprehensive Reviews in Food Science and Food Safety. 2011 Sep.
105. Chen L, Daniel RM, Coolbear T. Detection and impact of protease and lipase activities in milk and milk powders. Int Dairy J. 2003;13:255–75.
106. Samaržija D, Zamberlin Š, Pogačić T. Psychrotrophic bacteria and milk quality. Mljekarstvo. 2012;62(2):77–95.
107. Akkerman M, Johansen LB, Rauh V, Sørensen J, Larsen LB, Poulsen NA. Relationship between casein micelle size, protein composition and stability of UHT milk. Int Dairy J. 2021 Jan 1;112.
108. Liu Y, Guo R. pH-dependent structures and properties of casein micelles. Biophys Chem. 2008 Aug;136(2–3):67–73.
109. Sinaga H, Bansal N, Bhandari B. Effects of milk pH alteration on casein micelle size and gelation properties of milk. Int J Food Prop. 2017 Jan 2;20(1):179–97.
Received: August 22, 2024 / Accepted: November 1, 2024 / Published: December 15, 2024
Citation: Molina E, Camacho R, Mejía D, Marcial-Coba M. Proteolytic and Biofilm-forming Capacity of Bacteria Isolated from a UHT Milk Processing Plant. Bionatura Journal 2024; 1 (4) 8. http://dx.doi.org/10.70099/BJ/2024.01.04.8
Additional information Correspondence should be addressed to msmarcial@puce.edu.ec
Peer review information. Bionatura thanks anonymous reviewer(s) for their contribution to the peer review of this work using https://reviewerlocator.webofscience.com/
ISSN.3020-7886
All articles published by Bionatura Journal are made freely and permanently accessible online immediately upon publication, without subscription charges or registration barriers.
Publisher's Note: Bionatura Journal stays neutral concerning jurisdictional claims in published maps and institutional affiliations.
Copyright: © 2024 by the authors. They were submitted for possible open-access publication under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).