

Evaluación de Hongos Micorrízicos Arbusculares asociados con Miconia bracteolata Bonpl caso de estudio de Bosque Alto Andino, Ecuador
Evaluation of Arbuscular Mycorrhizal Fungi Associated with Miconia bracteolata Bonpl case study High Andean Forest, Ecuador

Marco Fabian Pilco-Pomagualli 1, Franklin Enrique Cargua-Catagña 2, Jorge Reyes Reyes 4,
Fernando Paredes-Arcos 5.
Fernando Paredes-Arcos 5.
1 Escuela Superior Politécnica de Chimborazo, Grupo de Investigación y Desarrollo para el Ambiente y Cambio Climático (GIDAC). Panamericana Sur km 1 1/2, Riobamba-Ecuador;
1 Escuela Superior Politécnica de Chimborazo, Grupo de Investigación y Desarrollo para el Ambiente y Cambio Climático (GIDAC). Panamericana Sur km 1 1/2, Riobamba-Ecuador;
1 Escuela Superior Politécnica de Chimborazo, Grupo de Investigación y Desarrollo para el Ambiente y Cambio Climático (GIDAC). Panamericana Sur km 1 1/2, Riobamba-Ecuador franklin.cargua@espoch.edu.ec .
2 Programa de Doctorado Internacional en Agricultura y Medioambiente para el Desarrollo, Universidad Santiago de Compostela, Escuela Politécnica Superior, Campus Universitario S/n, 27002-Lugo, España.
3 Facultad de Ingeniería/Ingeniería Ambiental, Universidad Nacional de Chimborazo, Ecuador-UNACH, Riobamba, Ecuador, carla.silva@unach.edu.ec .
4 Cuerpo Académico en Recursos Forestales, Facultad de Ciencias Agrícolas, Universidad Autónoma de Chiapas. Entronque Carretera costera y Estación Huehuetán; Huehuetán, Chiapas, México. C.P. 30670. jorge.reyes@unach.mx .
5 Programa de Estadística Aplicada, Universidad Politécnica Estatal del Carchi. Carchi-Ecuador; fernando.paredes@upec.edu.ec .
* Correspondencia: diego.damian@espoch.edu.ec; +593 99 817 2874, Diego Armando Damián.
RESUMEN
El objetivo de la investigación es identificar los hongos micorrízicos arbusculares (HMA) asociados a Miconia bracteolata Bonpl, determinando los parámetros ecológicos. La presente investigación centra su análisis en el sector de Guangra, ubicado en la zona de amortiguamiento del parque Nacional Sangay – Ecuador, en donde se empleó metodologías de vanguardia para la extracción y cuantificación de HMA. Se identificó 14 géneros de morfotipos de HMA asociadas a Miconia bracteolata Bonpl, la mayor población de espora encontradas corresponde a la parcela 03 (5637,95 esporas/100g de suelo); mientras que el valor inferior se presentó en la parcela 01 (3586,46 g/100g de suelo); el porcentaje de colonización promedio de 15.69, las cuales se desarrollando en un tipo de suelo con pH ligeramente acido que está asociado fuertemente a la supervivencia de las esporas de los HMA. Los parámetros ecológicos muestran un valor de importancia elevado en la parcela 03 en el género Glomus spp; un índice de H con diversidad media y un índice de diversidad de Simpson de alta dominancia. Estos resultados ayudaran a mejorar la capacidad de predecir distribuciones como el grado de intervención de los ecosistemas, ayudando a los esfuerzos para conservar la biodiversidad y funciones de los ecosistemas altoandinos.
Palabras clave: Andes Ecuatorianos, bosques de ceja andina, índices de diversidad florística, morfotipos de hongos micorrícicos.
ABSTRACT
The research aims to identify the arbuscular mycorrhizal fungi (AMF) associated with Miconia bracteolata Bonpl and determine the ecological parameters. The analysis focuses on the Guangra sector, located in the buffer zone of the Sangay National Park - Ecuador, using the most modern methods for the extraction and quantification of AMF. The highest spore populations were found in plot 03 (5637.95 spores/100 g soil), while the lowest was found in plot 01 (3586.46 spores/100 g soil), with a mean colonization percentage of 15.69, developed in a soil type with a slightly acidic pH, which is strongly associated with AMF spore survival. In plot 03, the genus Glomus spp. showed high importance for ecological parameters, medium diversity H-index, and high dominance Simpson's diversity index. These results can help to predict distributions and the degree of ecosystem disturbance, ultimately contributing to the conservation of biodiversity and functions in high Andean ecosystems.
Keywords: Ecuadorian Andes, Andean brow forests, floristic diversity indices, mycorrhizal fungal morphotypes.
INTRODUCCIÓN
El bosque nuboso andino denominado “bosque siempre verde de ceja andina” ubicado en el callejón interandino de los andes Ecuatorianos es reconocido como uno de los más importantes centros de biodiversidad florística del mundo, siendo considerados como hot spots dentro de hot spots1 , el cual se extiende en el Ecuador sobre los 3000 m.s.n.m., encontrándose en la zona de transición de bosque a páramo, volviéndole rico en diversidad florística; pero al mismo tiempo este tipo de ecosistema natural ha perdido posiblemente más de la mitad de su cobertura original por afectaciones antrópicas2 . Dentro de este ecosistema encontramos diferentes tipos de vegetación y una de las especies arbóreas más representativas es Miconia bracteolata Bonpl, que por lo general se encuentran en zonas montañosas aisladas de la urbanización y en terrenos con alta humedad. Su característica principal son sus hojas con venación acródroma en la que uno o más pares de venas laterales son muy desarrolladas, las cuales convergen desde la base hacia el ápice en forma de arco perteneciendo a la familia Melastomataceae3 .
Los HMA se asocian con más de 80% de las plantas terrestres de forma habitual y se producen a nivel mundial en la mayoría de los ecosistemas terrestres4 . El tipo y estado de la vegetación es un factor clave en la simbiosis entre el suelo y microorganismos edáficos como los hongos, nematodos y bacterias, que influyen directamente en los procesos de descomposición de la materia orgánica y el ciclo de nutrientes, por lo cual existe la necesidad de caracterizar las comunidades de microorganismos del suelo5 . En esta variada microfauna encontramos a los hongos micorrícicos arbusculares (HMA) que se asocian con más de 80% de las plantas terrestres de forma habitual y se producen a nivel mundial en la mayoría de los ecosistemas terrestres4 , regulados por factores climáticos en constante relación con el ambiente, también se encuentran en suelos de bajas y medias altitudes con altas tasas de mineralización de materia orgánica, colonizando raíces de múltiples plantas herbáceas y leñosas6 . Los HMA mediante sus micelios examinan el suelo logrando la transferencia de nutrientes a la planta por simbiosis a través de estructuras interradiculares especializadas (hifas) que constituye el cuerpo vegetativo del hongo, el cual cumplen funciones como: movilizar y transportar nutrientes principalmente carbono (C), nitrógeno (N) y fósforo (P), además minimiza el estrés hídrico, repara la agregación del suelo en procesos de erosión, brinda la protección de patógenos, ayuda en la estabilización del suelo, entre otros7 .
Los HMA crean estructuras especializadas (arbusculos – vesículas) por lo cual se les denomina “simbiontes obligados” los cuales utilizan la fotosíntesis del huésped para su crecimiento8 . La simbiosis entre la raíz de la planta y los HMA puede ser denominada como bio-inoculante para reducir la insuficiencia de nutrientes en la planta9 ,10 siendo amplio el impacto de las interacciones ecológicas de las comunidades fúngicas con las vegetales y la comunidad microbiana edáfica11 , esta asociación influye en los procesos de los ecosistemas para inducir una vasta variedad de respuestas al crecimiento con la planta que coexiste, un ejemplo de esto es la alta taza de descomposición de la hojarasca que tienen las plantas asociadas a HMA y estas influyen en el aporte de materia orgánica y la dinámica de C y N en el suelo12 .
Algunos HMA se benefician en mayor o menor grado dependiendo de las especies de plantas herbáceas que se relacionen, debido a que las asociaciones simbióticas con hongos difieren de las especies de plantas que están dentro del sotobosque, el cual se caracteriza por tener la mayor riqueza que los demás estratos del bosque13 . Además, existe variación en la absorción de los nutrientes de las plantas huésped de HMA debido al tipo de grupo funcional y su morfología radicular14 .
La diversidad de los HMA se estima sobre la base de la cantidad de esporas, siendo este método problemático porque la esporulación fúngica se rige por varios factores, un ejemplo de ello es la distribución de la mayoría de los taxones de los HMA que se ven alterados por las asociaciones con diferentes tipos de familias de plantas o distintas zonas climáticas15 ,16 . Sin embargo, las estimaciones actuales de la diversidad fúngica de los HMA se basan en gran medida de la morfología de las esporas como: forma, color, tamaño, grosor, tipo y naturaleza de las paredes que la constituyen, conexión hifal, ornamentaciones, entre otras17 .
El objetivo de presente estudio fue identificar los hongos micorrízicos asociados a Miconia bracteolata Bonpl en condiciones naturales, debido a que no existen estudios sobre el tipo anatómico de colonización que desarrolla esta especie, con el fin de conocer la ocurrencia, importancia y la diversidad de HMA en los Bosque Siempre Verde de Ceja Andina.
Fase de campo
Diseño y método de Muestreo
Se realizó una rejilla de puntos de 1 Km x 1 Km en el sitio de estudio, del cual se tomó al azar un conglomerado, donde se estableció tres parcelas en forma “L”, con una dimensión 60 m x 60 m., con una distancia entre parcelas de 250 m. De cada parcela se tomaron 10 muestras (arboles) las mismas que se homogenizaron hasta completar 1 kilogramo, la muestra abarco el sistema radicular para obtener suelo y raíces con micorrizas a una profundidad entre 0 a 20 cm (ver Figura 2).
Análisis en laboratorio
El análisis de muestras para determinar las micorrizas se realizó en la Universidad Nacional de Chimborazo, Facultad de Ingeniería, laboratorio de servicios ambientales (LSA-UNACH) y en el Laboratorio del Departamento de suelos de la Facultad de Recursos Naturales de la Escuela Superior Politécnica de Chimborazo.
Extracción y cuantificación de micorrizas arbusculares
Se separó las esporas del hongo, preparando una muestra de 100 g de suelo compuesto, aplicando peróxido de hidrogeno al 1.5% por una hora, se disgrego la muestra empleando tamices de 150 y 75 μm (se lavó), el tamizado de 150 y 75 μm se licuo a baja velocidad por 30 segundos y por 60 segundos a velocidad alta respectivamente, las muestras se recogieron por separado sobre un papel filtro en el interior de un embudo de Buchner para filtrarse al vacío, para dejarlas secar a temperatura ambiente. Posteriormente se pesó 1 g de suelo de cada tamiz, se colocó en tubos de centrifuga a los cuales se añadió 20 ml de H2O y 20 ml de sacarosa al 2M, centrifugándose por 5 minutos a 4000 r.p.m. concluido se lavó las esporas y se recogieron las muestras que quedaron sobre cada tamiz y se colocaron en cajas Petri cuadricuadas para cuantificar las esporas.
Caracterización Biológica
Se tomó muestras de raíces de la especie Miconia bracteolata Bonpl a una profundidad entre 0 y 20 cm, para medir la colonización se procedió a teñir las raíces empleando el siguiente proceso: Se pesó 1 g de raíces en un Erlenmeyer de 25 ml, luego se colocó KOH al 10% por una hora a 90 °C en baño maría. Posteriormente se lavó las raíces y se colocó en el Erlenmeyer con HCL 1N durante 15 min a temperatura ambiente, se escurrió el HCL y aplico Azul de Trypan al 0,05% en lactoglicerina y reposó por ½ hora, transcurrido este tiempo, la muestra pasó a baño María por 15 min a 90 °C, luego se lavó con abundante agua, se agregó lactiglicerina y se dejó en reposo por 24 horas, culminado el tiempo se procedió a evaluar la colonización utilizando el método de intersección de línea de cuadrícula16 .
Análisis fisicoquímico del suelo
Los parámetros fisicoquímicos del suelo analizados fueron: pH por medio del potenciométrico en suelo húmedo, materia orgánica (MO) por combustión húmeda (Walkley-Black) y para determinar elementos de interés por medio del Espectrómetro de Absorción Atómica AA-300.
Parámetros ecológicos
Índice de Valor de importancia
Permite comparar el peso ecológico de cada especie dentro de la población, el valor IVI indica igualdad o diferencia en su composición, estructura, calidad de sitio y dinámica19 .
Índice de diversidad de Simpson (D’)
Manifiesta la probabilidad de que dos individuos tomados al azar de una muestra sean de la misma especie. Está fuertemente influido por la importancia de las especies más dominantes20 .

D’ representa el Índice de Simpson; S es el número de especies, ni es el Número total de individuos presentes, i respecto al total de individuos (es decir la abundancia relativa de la especie i); y N es el número total de individuos de todas las especies.
Índice de Diversidad de Shannon (H’)
Mide el grado promedio de incertidumbre en predecir a que especie pertenecerá un individuo escogido al azar de una colección. Asume que los individuos son seleccionados al azar y que todas las especies están representadas en la muestra. Adquiere valores entre cero, cuando hay una sola especie, y el logaritmo de S, cuando todas las especies están representadas por el mismo número de individuos21 .

H' representa el Índice de Shannon-Wiener; S es el número de especies; y pi es la proporción de individuos de la especie i respecto al total de individuos (es decir la abundancia relativa de la especie i).
Análisis en estadístico
Mediante el Software Minitab se realizó un análisis estadístico descriptivo, además se realizó la prueba ANOVA con un nivel de significancia de 0.05, para comprobar que existe variación entre las variables, se procedió a realizar una prueba de TUKEY para determinar sus diferencias. Para determinar los Índices de Biodiversidad se utilizó el software Past 4.12b.
RESULTADOS
La recolección de muestras se desarrolló en el sector de Guangra, el cual se encuentra ubicado en la zona de amortiguamiento del parque Nacional Sangay, en la parroquia Achupallas, cantón Alausí, provincia de Chimborazo-Ecuador (Tabla 1 y Figura 1), con un tipo de cobertura corresponde a un bosque siempre verde de ceja andina el cual se distribuye desde los 3000 hasta los 3400 m.s.n.m, su vegetación está representada por: Miconia bracteolata, Myrsine andina y Aegiphila sp., con una superficie aproximada de 2240,92 ha, mismo que representa el 19% de la superficie total de la comunidad, con un suelo de tipo Andisol formado por ceniza volcánicas jóvenes, negras y gruesas18 , registrando una temperatura media anual de 8,3°C con temperaturas diarias que varían entre los 0 a 17 °C y humedad relativa del 83,6% (EMA_Jubal_2013).

Figura 1. Área de estudio.

Tabla 1. Ubicación geográfica de parcelas.
El diseño de muestro consistió en un conglomerado de 1000m x 1000m con 3 parcelas de muestreo en las cuales se seleccionó 10 unidades experimentales por parcela (ver Figura2).

Figura 2. Diseño de muestreo y recolección de muestras edáficas.
Evaluación de parámetros fisicoquímicos del suelo
Los parámetros fisicoquímicos nos muestran un contenido bajo de Nitrógeno, Calcio, Magnesio y Potasio, los cuales posiblemente se deba al proceso de lixiviación por la baja retención del suelo por la textura franco-arenosa ya que el agua es un conductor de movilidad de los elementos. Mientras que, el Fosforo y Materia Orgánica poseen contenidos altos debido a la densidad de vegetación en el bosque y un pH acido típico de los ecosistemas endémicos alto andinos (Tabla 2).

Niveles: Alto (A), Medio (M), Bajo (B), Acido (Ac)
Tabla 2. parámetros fisicoquímicos del suelo
Determinación de la población de espora en el suelo
Para el análisis de la colonización de esporas micorrícicas se identificaron valores asociados con la capacidad de colonización, además de los índices florísticos. Encontrando que la mayor población de espora en el suelo se identificó en la parcela 03 (5637,95 esporas/100g de suelo) y el valor más bajo se presentó en la parcela 01 (3586,46 g/100g de suelo), al realizar la prueba de ANOVA entre parcelas con un nivel de confianza de p≤ 0.05, se obtuvo un p valor de 0.015 determinando que existe diferencia entre la parcela 3 con las parcelas 2 y 1, las cuales presentan una correlación (Figura 3).

Figura 3. Distribución de la cantidad de Esporas/100 g de suelo por parcelas. Las medias que no comparten una letra son significativamente diferentes con nivel de confianza del 95%.
Se observó en campo que la parcela 1 y 2 se encuentran cerca de zonas de perturbación, afectando directamente a las poblaciones de hongos micorrícicos y la simbiosis planta–hongo.
El análisis de normalidad de los datos en las diferentes parcelas muestra una distribución normal (Figura 4) en las parcelas 01 y 03, con valores medios de 256,2 y 402.7 esporas/100g de suelo, lo que corresponde a que todos los valores se encuentran dentro de una normalidad con respecto a su media, mientras que la parcela 02 muestra una característica de valores alejados de la normalidad, pero los valores de p < 0,05 manifiesta que la variabilidad de colonización se presenta en las diferentes parcelas, dependiendo esta colonización de la densidad de especies asociadas a los HMA con Miconia bracteolata Bonpl.

Tabla 3. Morfotipos de esporas en 100 gramos de suelo por parcela.

Figura 4. Análisis de Normalidad de la parcela 01, 02 y 03
Porcentaje de colonización radicular en Miconia bracteolata Bonpl.
Las raíces de Miconia bracteolata Bonpl., evaluadas reportaron colonización por estructuras de hongos micorrícicos (hifa y arbusculos). Encontrando el mayor porcentaje de colonización en la parcela 03 con un valor de 17,14%, seguido de la parcela 02 con 15,11% y por último la parcela 01 con un valor de 14,84% (Figura 5), encontrando un valor p>0,05 demostrando que no existe diferencia entre parcelas.

Figura 5. Porcentaje de colonización radicular por parcela. Las medias que no comparten una letra son significativamente diferentes con nivel de confianza del 95%.
Determinación de parámetros ecológicos
Identificación de colonias en 100 g/suelo
Los resultados de los análisis micorrícicos en el suelo proveniente del Bosque de Ceja Andina alcanzaron un promedio de 4363,80 esporas en 100 gramos de suelo, este tipo de población es considerablemente alto, debido a las condiciones climáticas y abundante vegetación obteniendo 14 géneros de morfotipos de hongos micorrícicos como se observa en la Figura 6.
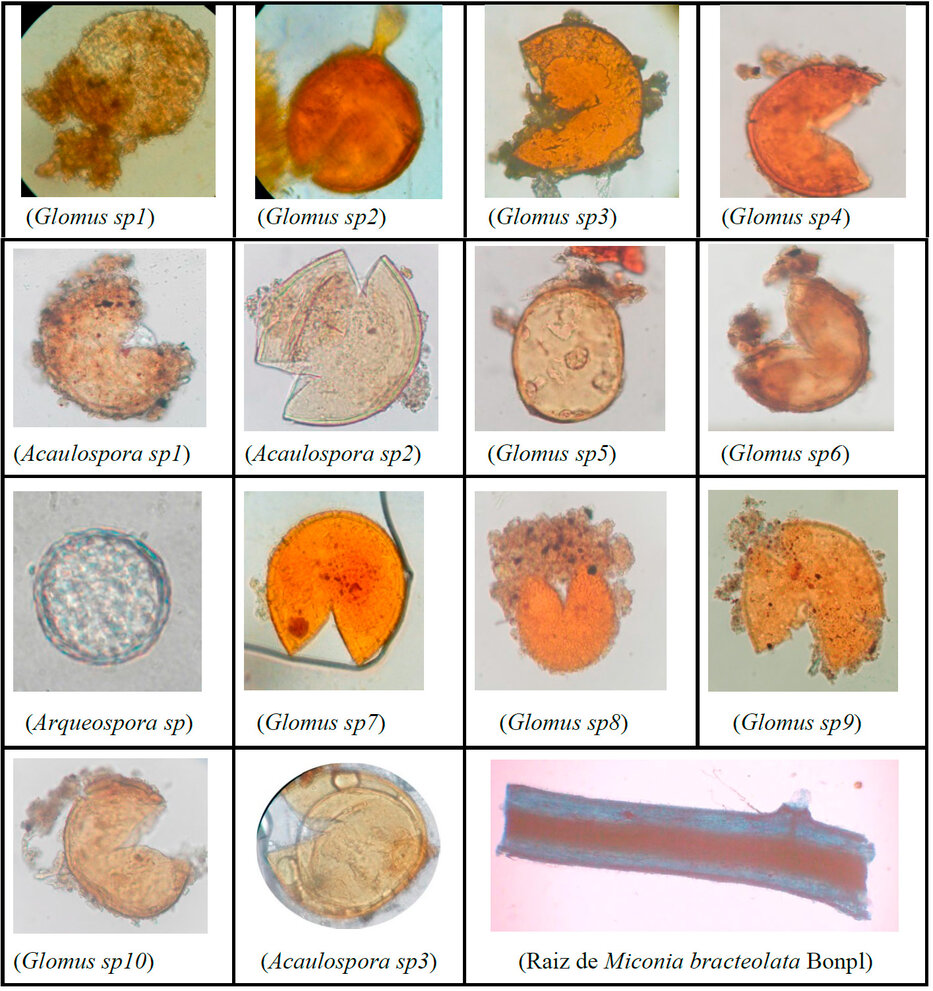
Figura 6. Morfotipos asociados a Miconia bracteolata Bonpl Aumento 100X y raíz 40X.
Valor de Importancia de géneros de esporas de hongos micorrícicos
La Tabla 4 revela la importancia que tienen las esporas micorrícicas dentro de la comunidad biológica del suelo en la parcela 01, el género Glomus sp3 que presenta el valor más alto con 12,68 y el género Acaulospora sp2 presenta el valor más bajo con 4,93. Mientras que, en la parcela 02 los géneros Glomus sp6 y Acaulospora sp3 presentan el valor más alto con 9, 20 y el género Glomus sp2 y Glomus sp5 el valor más bajo con 5,52. Finalmente, en la parcela 03 el género Glomus sp3 que presenta el valor más alto con 16,75 y el género Glomus sp10 presenta el valor más bajo con 4,44, a excepción del género Arqueospora sp., que solo encontramos en la parcela 03.

Tabla 4. Índice de Valor de importancia por parcela.
Índice de Diversidad de Shannon-Weaver (H)
El índice Shannon-Weaver expresa la uniformidad de los valores de importancia a través de todas las especies de la muestra, encontrando que las parcelas 02 y 03 registraron valores similares de diversidad (2,55), seguidos de la parcela 01 que presento un valor de 2,52; esto indica que la diversidad de hongos micorrícicos es media en la zona de estudio y esto podría deberse al grado de intervención antrópica en la parcela 01.
Índice de Diversidad de Simpson
La Figura 8, se muestra que las parcelas 01 y 03 registraron mayor Índice de Diversidad con un valor de 0,91, seguido por la parcela 02 que presentó un valor de 0,92; lo que indica que la diversidad de esporas de hongos micorrícicos presenta alta dominancia ya que los valores se acercan a 1. La baja riqueza de especies encontrada en la rizosfera de Miconia bracteolata bonpl podría reflejar limitaciones en los patrones de esporulación bajo condiciones de campo y los cultivos trampa pueden revelar un mayor nivel de HMA y diversidad en el suelo22 .

Figura 7. Índice de Diversidad de Simpson por parcela de estudio.
DISCUSIÓN
Análisis de micorrizas arbusculares, factores ambientales y edáficos
En el bosque siempre verde de ceja Andina, del sector Guangra, las poblaciones de HMA muestran variación, encontrando un total de 3587 en la parcela 01, 3867 parcela 02 y 5638 en la parcela 03 de esporas sobre 100g de suelo. Para Stover et al. en 201823 el tipo, magnitud y frecuencia de perturbación alteran la diversidad de los HMA, reduciéndose a la mitad el número de especies de HMA como se muestra en la Tabla 3. Mientras que la densidad de esporas de los hongos micorrícicos nativos podría estar influenciada por la clase textural del suelo franco-arenoso predominante en la zona de estudio, debido a que los suelos con textura arenosos no favorecen la simbiosis micorrícica que pueda establecerse con diversos hospederos, mientras que en suelos arcillosos incrementan favorablemente la simbiosis y la densidad de los hongos micorrícicos, por la relación que existe entre la textura y la disponibilidad de humedad del suelo 24 . El mayor o menor número de esporas presente en cada tipo de suelo, no necesariamente indica que se da una mayor infección de la raíz por el mismo organismo, sino que depende de la relación hongo-hospedero y esta simbiosis entre los HMA, además que el huésped ofrezca protección conocida como “resistencia inducida por micorrizas” los cuales son fundamentales y actúan de protección de algunos patógenos que pueden ser nocivos y alterar el ecosistema. Sin embargo, se existe la posibilidad que la incidencia de infección aumente con relación al número de esporas en el suelo, indicándonos que el nivel de colonización radicular también depende si la especie Miconia bracteolata Bonpl, es altamente micorrizable, y que los géneros de hongos encontrados en el suelo no sean infectivos o efectivos con esta especie forestal25 .
En este contexto la presencia de HMA también dependen del tipo de vegetación, estas asociaciones con especies arbóreas de madera dura se asocian a HMA y algunas especies arbóreas de coníferas se asocian a ectomicorrizas, debido a que los HMA son menos específicos de la especie que los hongos ectomicorrícicos26 . La dependencia de los HMA hace que algunas especies arbóreas tengan y estén asociadas a una red de micelios que a su vez puedan actuar como reservorios de hongos, algo similar ocurre en este trabajo de investigación, en el cual existe mayor presencia de Miconia bracteolata Bonpl. pero también influye en la presencia o ausencia de especies de HMA en cada área lo cual determina la composición de la comunidad. Demostrando la capacidad que tienen los HMA de permanecer en el suelo asociado a plantas huésped y la variabilidad en sus estructuras, ayudándonos a predecir disturbios en el ecosistema lo cual aporta con el objetivo de conservar la biodiversidad y funciones del ecosistema y este importante grupo de hongos5,27 .
En la presente investigación se encontró un contenido bajo de Nitrógeno, Calcio, Magnesio y Potasio, y altos para Fosforo (49 mg/L). Los altos nivel de fosforo, mejoran la comunidad fúngica aumentando la diversidad y riquezas de los HMA28,29 . Mientras que Moína-Quimí et al. mencionan que el porcentaje de infección micorrícica en las plantas está fuertemente determinado por el grado de acidez del suelo y la supervivencia de las esporas de los HMA30 , encontrando en nuestro caso de estudio un tipo de suelo con pH ligeramente acido. Algunos autores sugieren que el porcentaje de colonización depende del grado de nutrientes presentes en los suelos, los cuales disminuyen a medida que la altitud aumenta debido a que la temperatura disminuye lo que ocasiona que el ciclo de nutrientes se vuelva lento4 . Para Johnson et al., los suelos pobres o infértiles fortalecen a la asociación con las micorrizas, debido a que los hospederos generan más simbiosis con estructuras que les favorecen a la adquisición del recurso limitante31 .
Parámetros ecológicos
Los resultados de la presente investigación se corroboran con el estudio realizado por32 el cual menciona que el 83% de la comunidad global de HMA son de los géneros Acaulospora y Glomus, su estudio fue realizado en un ecosistema similar al nuestro de Bosque. Autores como Bainard et al. en 201133 y Chiquito et al. en 201834 mencionan que la alta diversidad entre sitios puede ser el resultado de múltiples mecanismos, que no necesitan ser mutuamente excluyentes; una combinación de limitación de dispersión, efectos del huésped de la planta y variación en otras condiciones ambientales como la temperatura del suelo y la humedad, podría resultar en un amplio abanico de diversas composiciones, observado entre sitios altas tasas de variación espacial. La restricción en la composición de especies arbóreas y hongos en su mayoría pertenecientes a Glomus spp. fue reportado en las parcelas que se sitúan al borde del bosque, donde existe alteración de su ecosistema y baja diversidad vegetativa. Mostrando la importancia de HMA en los suelos forestales, la cual radica en el aprovechamiento de ciertos elementos químicos y especialmente en la transferencia de nutrientes a las plantas35 .
La diversidad que se muestra en la zona de estudio está reflejada en las diferentes condiciones y características que pueden presentar los ecosistemas altoandinos. Para Caicedo Rosero et al. en 201736 y Lazo et al. en 200737 , las condiciones climáticas intuyen en el desarrollo de las plantas, que a su vez determinan el crecimiento de la fauna y microfauna, los cambios en el número de especies e individuos de una especie a otra es diferente en los diversos periodos de precipitación en el hábitat donde se desarrollan los HMA y su presencia dependen de la temperatura, humedad del suelo, entre otros. Esta alteración en la diversidad fúngica puede limitar la nutrición de las plantas, ocasionado la desestabilidad en el ecosistema38 ,
Estrada et al. en 201339 menciona que la baja riqueza de HMA, se puede deber a que las especies pueden estar más adaptadas para ajustar los patrones de esporulación a las condiciones de estrés ambiental. Los HMA en su mayoría pertenecientes a Glomus spp. tienen mayor capacidad de esporulación en ecosistemas que presentan condiciones extremas en el tipo de suelo y clima. Sin embargo, Longo et al. En 201440 muestra que los cambios en las comunidades de esporas de HMA podría ser un efecto directo de cambios en la vegetación dentro de ecosistemas alterados. Por otro lado, Camenzind & Rillig. en 201341 menciona que los suelos altamente orgánicos están ligados íntimamente con el desarrollo de HMA ya que ellos forman un micelio de gran tamaño dentro de la capa orgánica de un suelo, el cual se encuentra asociado íntimamente con la descomposición del material; mientras que Montero Sommerfeld et al., en 201342 indica en sus estudios que la comunidad de HMA puede variar significativamente de acuerdo a la profundidad y perfiles del suelo relacionados con la composición química y estructura; así también el pH del suelo influye en la distribución espacial de las diferentes especies de HMA, es por ellos algunas familias son menos abundantes43,44 .
Análisis reflexivo de las metodologías y resultados en la investigación de HMA asociados a Miconia bracteolata.
El muestreo de este estudio consiste en tres parcelas dentro de las cuales se seleccionaron 10 árboles de Miconia bracteolata de manera aleatoria por parcela. Dalli et al. en 2020 realizaron un muestreo similar en el cual seleccionaron 10 individuos de la especie Ceratonia siliqua L47. Mientras tanto, Herrmann et al. en 2016 seleccionaron 9 árboles de Hevea brasiliensis para su estudio54. Este tipo de muestreo puede ser realizado en un solo sitio, como en nuestro estudio y en los realizados por Erazo, Manzano y Patiño en 2020 en el bosque Llucud y Palictahua52, que también se enfocaron en caracterizar las interacciones de los HMA con una especie específica. Este enfoque proporciona información valiosa sobre la actividad de los HMA en un contexto específico. Por otro lado, existen estudios que se llevaron a cabo en múltiples localidades, como el de Gao et al. en 202155, quienes analizaron la diversidad de HMA en 11 ciudades de la isla de Hainan. Dalli et al. en 2020 también realizaron su levantamiento en tres localidades diferentes de Argelia47. Herrmann et al. en 2016 seleccionaron árboles de Hevea brasiliensis en diversas localidades de Tailandia54, y Mahdhi et al. en 2019 estudiaron tres especies de acacia en diferentes sitios del suroeste de Arabia Saudita46.Además, nuestra investigación se centró exclusivamente en los HMA asociados a Miconia bracteolata Bonpl, la especie más representativa de este bosque. Esto proporciona información específica sobre la simbiosis micorrícica de esta planta en particular. Sin embargo, este estudio establece la línea base para investigar las asociaciones de HMA con una variedad más amplia de especies vegetales, ofreciendo una visión holística de la comunidad micorrícica y su variabilidad en este ecosistema46,47.
El estudio se basó en la identificación de HMA mediante la morfología de las esporas, una técnica comúnmente utilizada debido a su bajo costo y facilidad de aplicación. Esta metodología es ampliamente empleada en estudios de diversidad de HMA en diversos ecosistemas debido a su simplicidad y a la infraestructura mínima requerida46-51. Esta metodología es ampliamente empleada en estudios de diversidad de HMA en diversos ecosistemas debido a su simplicidad y a la infraestructura mínima requerida47. La identificación morfológica permite la clasificación de esporas basándose en características como el tamaño, la forma, el color y la estructura de la pared, lo cual ha sido efectivo en estudios previos para diferenciar diversas especies de HMA en distintos contextos ambientales46-51,56. Sin embargo, aunque la morfología de esporas es una herramienta útil es recomendable complementar esta metodología con análisis moleculares56,57. Técnicas como la reacción en cadena de la polimerasa (PCR) y la secuenciación de ADN han demostrado ser efectivas para confirmar la identidad de las especies de HMA y proporcionar una mayor resolución taxonómica57,58.
La combinación de técnicas morfológicas y moleculares ofrece una visión más completa de la diversidad y estructura de las comunidades de HMA 56-58. En estudios previos, la integración de ambas metodologías ha permitido la identificación de una mayor diversidad de especies y ha proporcionado información más detallada sobre las relaciones filogenéticas entre las diferentes especies de HMA57. Por ejemplo, Gao et al. 2021 utilizaron tanto la identificación morfológica como molecular para caracterizar la diversidad de HMA en plantaciones de Hevea brasiliensis en la isla de Hainan, China, revelando una compleja comunidad de HMA influenciada por factores edáficos y la edad de la plantación55.
Además, la investigación realizada por Herrmann et al. en 2016 en Tailandia también utilizó ambas metodologías para estudiar la diversidad de HMA en plantaciones de Hevea brasiliensis, encontrando que la diversidad y abundancia de HMA varían significativamente entre localidades y están correlacionadas con las condiciones del suelo y el manejo agrícola54. De manera similar, Mahdhi et al. en 2019 llevaron a cabo estudios en tres sitios diferentes del suroeste de Arabia Saudita, combinando análisis morfológicos y moleculares para caracterizar las comunidades de HMA asociadas con especies de acacia, demostrando la importancia de los HMA en la sostenibilidad de estos ecosistemas áridos46.La identificación precisa de HMA es crucial para entender sus roles ecológicos y su contribución a la salud del suelo y la productividad vegetal58. Los HMA son conocidos por mejorar la absorción de nutrientes, especialmente fósforo, y por aumentar la resistencia de las plantas a condiciones de estrés abiótico como la sequía y la salinidad 56,57. Por lo tanto, una comprensión detallada de las comunidades de HMA puede informar prácticas de manejo sostenible y estrategias de reforestación, particularmente en ecosistemas degradados o bajo presión ambiental58.
CONCLUSIONES
Se identificaron 14 géneros de morfotipos de MHA asociadas a Miconia bracteolata Bonpl, la mayor población de espora encontradas en el suelo correspondió a la parcela 03 (5637,95 esporas/100g de suelo), mientras que el valor inferior se presentó en la parcela 01 (3586,46 g/100g de suelo). Se observó en campo que la parcela 01 se encuentra cerca de zonas de perturbación, afectando directamente a las poblaciones de hongos micorrízicos y la simbiosis planta-hongo; el porcentaje de colonización promedio fue de 15.69 las cuales se desarrollaron en un tipo de suelo con un pH ligeramente acido que está asociado fuertemente a la supervivencia de las esporas de los HMA.
Los parámetros ecológicos muestran un valor de importancia elevado en la parcela 03 en el género Glomus spp, un índice de H con diversidad media y un índice de diversidad de Simpson de alta dominancia. Estos datos ayudaran a mejorar la capacidad de predecir distribuciones como el grado de intervención de los ecosistemas, ayudando a los esfuerzos para conservar la biodiversidad y funciones de los ecosistemas altoandinos.
Los parámetros ecológicos muestran un valor de importancia elevado en la parcela 03 en el género Glomus spp, un índice de H con diversidad media y un índice de diversidad de Simpson de alta dominancia. Estos datos ayudaran a mejorar la capacidad de predecir distribuciones como el grado de intervención de los ecosistemas, ayudando a los esfuerzos para conservar la biodiversidad y funciones de los ecosistemas altoandinos.
Contribuciones de los autores: Conceptualización, D.D.-C. y .M.P.-P.; metodología, D.D.-C.; software, D.D.-C. y J.R.R; validación, D.D.-C., M.P.-P., F.C.-C. y C.S.-P.; análisis formal, .D.D.-C. y F.C.-C; investigación, .D.D.-C; recursos, D.D.-C; curación de datos, F.P.-A. y J.R.R; redacción: preparación del borrador original, D.D.-C.; redacción: revisión y edición, D.D.-C., C.S.-P., M.P.-P., Y F.P.-A.; visualización, .D.D.-C; supervisión, .D.D.-C.; administración de proyectos, .D.D.-C.; adquisición de financiación, D.D.-C. Todos los autores han leído y aceptado la publicación de este manuscrito.
Financiamiento: Está investigación no recibió financiamiento externo.
Agradecimientos: Al Grupo de Investigación y Desarrollo para el Ambiente y Cambio Climático, de la Escuela Superior Politécnica de Chimborazo “GIDAC-ESPOCH”. A la Universidad Nacional de Chimborazo, por el uso de las instalaciones y asesoría en la carrera de Ambiental de la Facultad de Ingeniería. A la Doctora Norma Erazo por el apoyo en las metodologías de la fase de campo y laboratorio. Y todas las personas de la parroquia Achupallas por el apoyo brindado en el desarrollo de esta investigación.
Conflictos de intereses: Los autores declaran no tener conflicto de intereses
REFERENCIAS
1. Luna-Florin AD, Nole-Nole DA, Rodríguez-Caballero E, et al. Ecological characterization of the flora in Reserva Ecológica Arenillas, Ecuador. Applied Sciences 2022; 12(17):8656; doi: 10.3390/app12178656.
2. Zuluaga S, Vargas FH, Aráoz R, et al. Main aerial top predator of the andean montane forest copes with fragmentation, but may be paying a high cost. Glob Ecol Conserv 2022; 37:02174; doi:10.1016/j.gecco.2022.e02174.
3. Goldenberg R, Penneys DS, Almeda F, et al. Phylogeny of Miconia (Melastomataceae): Patterns of stamen diversification in a megadiverse neotropical genus. Int J Plant Sci 2008;169(7):963–979; doi: 10.1086/589697.
4. Camenzind T, Homeier J, Dietrich K, et al. Opposing effects of nitrogen versus phosphorus additions on my-corrhizal fungal abundance along an elevational gradient in tropical montane forests. Soil Biol Biochem 2016; 94:37– 47; doi: 10.1016/j.soilbio.2015.11.011.
5. Stefanowicz AM, Kapusta P, Stanek M, et al. Herbaceous plant species and their combinations positively affect soil microorganisms and processes and modify soil physicochemical properties in a mesocosm experi-ment. For Ecol Manage 2023; 532:120826; doi: 10.1016/j.foreco.2023.120826.
6. Huang B, Zhang L, Cao Y, et al. Effects of land-use type on soil organic carbon and carbon pool management index through arbuscular mycorrhizal fungi pathways. Glob Ecol Conserv 2023; 43:02432; doi: 10.1016/j.gecco.2023.e02432.
7. Carron AI, Garibaldi LA, Marquez S, et al. The soil fungal community of native woodland in Andean Pata-gonian forest: A case study considering experimental forest management and seasonal effects. For Ecol Manage 2020; 461:117955; doi: 10.1016/J.FORECO.2020.117955.
8. Dey M, Ghosh S. Arbuscular mycorrhizae in plant immunity and crop pathogen control. Rhizosphere 2022; 22:100524; doi: 10.1016/j.rhisph.2022.100524.
9. Lozano Sánchez JD, Armbrecht I, Montoya Lerma J. Hongos formadores de micorrizas arbusculares y su efecto sobre la estructura de los suelos en fincas con manejos agroecológicos e intensivos. Acta Agron 2015;64(4); doi: 10.15446/acag.v64n4.46045.
10. Leocadio C, Álvarez-López N, Barrios A, et al. Soil culturable microbial diversity in an undisturbed montane cloud forest of Oaxaca, Mexico. Rev Mex Biodivers 2023; 94:944980; doi: 10.22201/ib.20078706e.2023.94.4980.
11. Wang Y, Zhang W, LI C, et al. Long-term nitrogen and/or phosphorus additions lead to a shift in the diversity of soil arbuscular mycorrhizal fungi and improve rainfed crop yield on the Chinese Loess Plateau. Pe-dosphere 2023; doi: 10.1016/j.pedsph.2023.01.009.
12. Fang M, Liang M, Liu X, et al. Abundance of saprotrophic fungi determines decomposition rates of leaf litter from arbuscular mycorrhizal and ectomycorrhizal trees in a subtropical forest. Soil Biol Biochem 2020; 149:107966; doi: 10.1016/j.soilbio.2020.107966.
13. Zubek S, Rożek K, Stefanowicz AM, et al. The impact of beech and riparian forest herbaceous plant species with contrasting traits on arbuscular mycorrhizal fungi abundance and diversity. For Ecol Manage 2021; 492:119245; doi: 10.1016/j.foreco.2021.119245.
14. Yang X, Ma Y, Zhang J, et al. How arbuscular mycorrhizal fungi drives herbaceous plants’ C: N: P stoichiom-etry? A meta-analysis. Science of The Total Environment 2023; 862:160807; doi: 10.1016/j.scitotenv.2022.160807.
15. Segovia-Cruz JA, Souza V, Mercado-Flores Y, et al. Microbial diversity of the Tecozautla geyser, Mexico. Rev Mex Biodivers 2023; 94:945131; doi: 10.22201/ib.20078706e.2023.94.5131.
16. Lee E-H, Eo J-K, Ka K-H, et al. Diversity of Arbuscular Mycorrhizal Fungi and Their Roles in Ecosystems. Mycobiology 2013;41(3):121–125; doi: 10.5941/MYCO.2013.41.3.121.
17. Sharma MP, Buyer JS. Comparison of biochemical and microscopic methods for quantification of arbuscular mycorrhizal fungi in soil and roots. Applied Soil Ecology 2015; 95:86–89; doi: 10.1016/j.apsoil.2015.06.001.
18. García VJ, Márquez CO, Isenhart TM, et al. Evaluating the conservation state of the páramo ecosystem: An object-based image analysis and CART algorithm approach for central Ecuador. Heliyon 2019;5(10); doi: 10.1016/j.heliyon.2019.e02701.
19. Alvis Gordo JF. Análisis estructural de un bosque natural localizado en zona rural del municipio de Popayán. Biotecnología en el sector Agropecuario y Agroindustrial 2009;7(1):115–122.
20. Simpson EH. Measurement of Diversity. Nature 1949 ;163(4148):688–688; doi: 10.1038/163688a0.
21. Daly A, Baetens J, De Baets B. Ecological Diversity: Measuring the Unmeasurable. Mathematics 2018; 6(7):119; doi: 10.3390/math6070119.
22. Chaudhary VB, Holland EP, Charman-Anderson S, et al. What are mycorrhizal traits? Trends. Ecol Evol 2022; 37(7):573–581; doi: 10.1016/J.TREE.2022.04.003.
23. Stover HJ, Naeth MA, Boldt-Burisch K. Soil disturbance changes arbuscular mycorrhizal fungi richness and composition in a fescue grassland in Alberta Canada. Applied Soil Ecology 2018; 131:29–37; doi: 10.1016/j.apsoil.2018.07.008.
24. Moebius-Clune DJ, Moebius-Clune BN, van Es HM, et al. Arbuscular mycorrhizal fungi associated with a single agronomic plant host across the landscape: Community differentiation along a soil textural gradient. Soil Biol Biochem 2013; 64:191–199; doi: 10.1016/j.soilbio.2012.12.014.
25. Nguvo KJ, Gao X. Weapons hidden underneath: bio-control agents and their potentials to activate plant in-duced systemic resistance in controlling crop Fusarium diseases. Journal of Plant Diseases and Protection 2019;126(3):177–190; doi: 10.1007/s41348-019-00222-y.
26. Seiwa K, Negishi Y, Eto Y, et al. Successful seedling establishment of arbuscular mycorrhizal-compared to ectomycorrhizal-associated hardwoods in arbuscular cedar plantations. For Ecol Manage 2020; 468:118155; doi: 10.1016/J.FORECO.2020.118155.
27. Xiao D, Chen Y, He X, et al. Temperature and precipitation significantly influence the interactions between arbuscular mycorrhizal fungi and diazotrophs in karst ecosystems. For Ecol Manage 2021; 497:119464; doi: 10.1016/j.foreco.2021.119464.
28. Liu Z, Bai J, Qin H, et al. Application of rice straw and horse manure coameliorated soil arbuscular mycor-rhizal fungal community: Impacts on structure and diversity in a degraded field in Eastern China. Land Degrad Dev 2021; 32(8):2595–2605; doi: 10.1002/ldr.3927.
29. Qin Z, Zhang H, Feng G, et al. Soil phosphorus availability modifies the relationship between AM fungal diversity and mycorrhizal benefits to maize in an agricultural soil. Soil Biol Biochem 2020; 144:107790; doi: 10.1016/j.soilbio.2020.107790.
30. Moína-Quimí E, Oviedo-Anchundia R, Nieto-Barciona S, et al. Evaluación de los Hongos Micorrízicos Ar-busculares de zonas del trópico húmedo del Ecuador. Bionatura 2018; 3(1); doi: 10.21931/RB/2018.03.01.9.
31. Johnson NC, Wilson GWT, Wilson JA, et al. Mycorrhizal phenotypes and the Law of the Minimum. New Phytologist 2015;205(4):1473–1484; doi: 10.1111/nph.13172.
32. Haug I, Setaro S, Suárez JP. Arbuscular Mycorrhizal Fungi and Ectomycorrhizas in the Andean Cloud Forest of South Ecuador. The Andean Cloud Forest 2021; 111–129; doi: 10.1007/978-3-030-57344-7_6/FIGURES/8.
33. Bainard LD, Klironomos JN, Gordon AM. Arbuscular mycorrhizal fungi in tree-based intercropping systems: A review of their abundance and diversity. Pedobiologia (Jena) 2011; 54(2):57–61; doi: 10.1016/j.pedobi.2010.11.001.
34. Chiquito Noboa I, Naranjo Morán J, Barcos-Arias M. Presencia de hongos micorrízicos arbusculares (HMA) en especies representativas del bosque seco tropical del litoral ecuatoriano. Bionatura 2018;3(1); doi: 10.21931/RB/2018.03.01.7.
35. Chen J, Heikkinen J, Hobbie EA, et al. Strategies of carbon and nitrogen acquisition by saprotrophic and ec-tomycorrhizal fungi in Finnish boreal Picea abies-dominated forests. Fungal Biol 2019; 123(6):456–464; doi: 10.1016/j.funbio.2019.03.005.
36. Caicedo Rosero DM, Benavides Rosales HR, Carvajal Pérez LA, et al. Población de macrofauna en sistemas silvopastoriles dedicados a la producción lechera: análisis preliminar. La Granja 2017; 27(1):77–85; doi: 10.17163/lgr.n27.2018.06.
37. Lazo JA, Valdés NV, Sampaio RA, et al. Diversidad zoológica asociada a un silvopastoreo leucaena-guinea con diferentes edades de establecimiento. Pesqui Agropecu Bras 2007; 42(12):1667–1674; doi: 10.1590/S0100-204X2007001200001.
38. Zhao Z, Wei X, Wang X, et al. Concentration and mineralization of organic carbon in forest soils along a climatic gradient. For Ecol Manage 2019; 432:246–255; doi: 10.1016/J.FORECO.2018.09.026.
39. Estrada B, Beltrán-Hermoso M, Palenzuela J, et al. Diversity of arbuscular mycorrhizal fungi in the rhizo-sphere of Asteriscus maritimus (L.) Less., a representative plant species in arid and saline Mediterranean ecosystems. J Arid Environ 2013; 97:170–175; doi: 10.1016/j.jaridenv.2013.05.019.
40. Longo S, Nouhra E, Goto BT, et al. Effects of fire on arbuscular mycorrhizal fungi in the Mountain Chaco Forest. For Ecol Manage 2014; 315:86–94; doi: 10.1016/j.foreco.2013.12.027.
41. Camenzind T, Rillig MC. Extraradical arbuscular mycorrhizal fungal hyphae in an organic tropical montane forest soil. Soil Biol Biochem 2013; 64:96–102; doi: 10.1016/j.soilbio.2013.04.011.
42. Montero Sommerfeld H, Díaz LM, Alvarez M, et al. High winter diversity of arbuscular mycorrhizal fungal communities in shallow and deep grassland soils. Soil Biol Biochem 2013; 65:236–244; doi: 10.1016/j.soilbio.2013.06.002.
43. Kohout P, Sudová R, Janoušková M, et al. Comparison of commonly used primer sets for evaluating arbus-cular mycorrhizal fungal communities: Is there a universal solution?. Soil Biol Biochem 2014; 68:482–493; doi: 10.1016/J.SOILBIO.2013.08.027.
44. Püschel D, Kolaříková Z, Šmilauer P, et al. Survival and long-term infectivity of arbuscular mycorrhizal fun-gi in peat-based substrates stored under different temperature regimes. Applied Soil Ecology 2019; 140:98–107; doi: 10.1016/J.APSOIL.2019.04.020.
45. Wang J, Wang GG, Zhang B, Yuan Z, Fu Z, Yuan Y, Zhu L, Ma S, Zhang J. Arbuscular Mycorrhizal Fungi Associated with Tree Species in a Planted Forest of Eastern China. Forests. 2019; 10(5):424. https://doi.org/10.3390/f10050424
46. Mahdhi M, Tounekti T, Abada E, Al‐Faifi Z, Khemira H. Diversity of arbuscular mycorrhizal fungi associated with acacia trees in southwestern Saudi Arabia. J Basic Microbiol. 2020 Apr 16;60(4):322–30. https://doi.org/10.1002/jobm.201900471
47. Dalli Y; Yahia N; Bekki A. Diversity of arbuscular mycorrhizal fungi associated with carob trees (Ceratonia Siliqua L.) in Western Algeria. Plant Cell Biotechnology and Molecular Biology. 2020, p. 180-193.
48. Lugo M.A, Ontivero R.E; Iriarte H.J; Yelikbayev, B.; Pagano, M.C. The Diversity of Arbuscular Mycorrhizal Fungi and Their Associations in South America: A Case Study of Argentinean and Brazilian Cattle Raising Productive Ecosystems: A Review. Diversity 2023, 15, 1006. https://doi.org/10.3390/d15091006
49. Jie W, Baiyan C, and Jingping G. Molecular detection and community analysis of arbuscular mycorrhizal fungi in the rhizosphere of Phellodendron amurense. Annals of microbiology. 2012: 1769-1777.
50. Marleau, J., Dalpé, Y., St-Arnaud, M., & Hijri, M. Spore development and nuclear inheritance in arbuscular mycorrhizal fungi. BMC evolutionary biology. 2011. 11, 1-11.
51. Boyno G, Demir S, Rezaee Danesh Y, Durak ED, Çevik R, Farda B, Djebaili R, Pellegrini M. A New Technique for the Extraction of Arbuscular Mycorrhizae Fungal Spores from Rhizosphere. Journal of Fungi. 2023; 9(8):845. https://doi.org/10.3390/jof9080845
52. Erazo N, Manzano J and Patiño B. Caracterización Molecular de la Diversidad Fúngica de los Bosques Llucud y Palictahua: Potencialidades en Control Biológico/Molecular Characterization of Diversity Fungic of the Llucud and Palictahua Forests: Potential in Biological Control. KnE Engineering. 2020. 313-328.
53. Singavarapu B, ul Haq H, Darnstaedt F, Nawaz A, Beugnon R, Cesarz S, et al. Influence of tree mycorrhizal type, tree species identity, and diversity on forest root‐associated mycobiomes. New Phytologist. 2024 May 24;242(4):1691–703. https://doi.org/10.1111/nph.19722
54. Herrmann L, Lesueur D, Bräu L, Davison J, Jairus T, Robain H, et al. Diversity of root-associated arbuscular mycorrhizal fungal communities in a rubber tree plantation chronosequence in Northeast Thailand. Mycorrhiza. 2016 Nov 23;26(8):863–77. https://doi.org/10.1007/s00572-016-0720-5
55. Gao X, Lv J, Guo C, Hu A, Wu X, Li Z. Species Diversity of Arbuscular Mycorrhizal Fungi in the Rhizosphere of Hevea brasiliensis in Hainan Island, China. Phyton (B Aires). 2021;90(1):179–92
56. Zhang S, Luo P, Yang J, Irfan M, Dai J, An N, et al. Responses of Arbuscular Mycorrhizal Fungi Diversity and Community to 41-Year Rotation Fertilization in Brown Soil Region of Northeast China. Front Microbiol. 2021 Oct 11;12. https://doi.org/10.3389/fmicb.2021.742651
57. Zhang M, Shi Z, Yang M, Lu S, Cao L, Wang X. Molecular Diversity and Distribution of Arbuscular Mycorrhizal Fungi at Different Elevations in Mt. Taibai of Qinling Mountain. Front Microbiol. 2021 Mar 4;12. https://doi.org/10.3389/fmicb.2021.609386
58. Cheng Z, Wu S, Du J, Liu Y, Sui X, Yang L. Reduced Arbuscular Mycorrhizal Fungi (AMF) Diversity in Light and Moderate Fire Sites in Taiga Forests, Northeast China. Microorganisms. 2023 Jul 19;11(7):1836. https://doi.org/10.3390/microorganisms11071836
Received: 8 December 2023/ Accepted: 20 July 2024 / Published: 15 September 2024
Citation: Damián-Carrión D A, Silva-Padilla C F, Pilco-Pomagualli M F, Cargua-Catagña F E, Reyes, Fernando Paredes-Arcos J R.Evaluación de Hongos Micorrízicos Arbusculares asociados con Miconia bracteolata Bonpl caso de estudio de Bosque Alto Andino, Ecuador.Bionatura Journal 2024; 1 (3) 10. http://dx.doi.org/10.70099/BJ/2024.01.03.10
Correspondence should be addressed to diego.damian@espoch.edu.ec
Peer review information. Bionatura thanks anonymous reviewer(s) for their contribution to the peer review of this work using https://reviewerlocator.webofscience.com/
ISSN. 3020-7886
All articles published by Bionatura Journal are made freely and permanently accessible online immediately upon publication, without subscription charges or registration barriers.
Publisher's Note: Bionatura Journal stays neutral concerning jurisdictional claims in published maps and institutional affiliations.
Copyright: © 2024 by the authors. They were submitted for possible open-access publication under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).